Jaltomata chihuahuensis (Bitter) Mione & Bye |
Mexico |
revised 14 Jan 2016 |
|
The information on this page may be cited as a communication with
professor Thomas Mione, Central Connecticut State University, Biology Department, Copernicus Hall, 1615 Stanley Street, New Britain, CT 06050-4010 USA |
Novon 6: 78-81. 1996. |
Link to Key to the Jaltomata species of Mexico |
|
|
|
 |
Figure 1. Side view of flower of Jaltomata chihuahensis (all anthers dehisced), and flower bud (not yet open) on right.
Photo by Thomas Mione, Mione 390 grown for study in the United States. |
 |
Figure 2. Flowers of Jaltomata chihuahensis, pistillate phase on left, hermaphroditic phase on right.
Units are mm. Photo by Thomas Mione, Mione 390 grown for study in the United States. |
 |
| Figure 3. Flower of Jaltomata chihuahensis (all five anthers dehisced). Photo by Thomas Mione, Mione 390 grown for study in the United States. |
 |
Figure 4. Back of flower of Jaltomata chihuahensis. Darker green is the calyx, lighter green is the corolla.
Units are mm. Photo by Thomas Mione, Mione 390 grown for study in the United States. |
 |
Figure 5. Fruits (green when ripe) and funnel-shaped calyx of Jaltomata chihuahensis.
Units are mm. Photo by Thomas Mione, Mione 390 grown for study in the United States. |
 |
Figure 6. Ripe fruits of Jaltomata chihuahensis fall free of the calyx.
At left is the calyx/pedicel without a fruit because the fruit has fallen.
Numbered units are cm. Photo by Thomas Mione, Mione 390 grown for study in the United States. |
 |
 |
Figure 7. Flower, units on ruler are mm; photographed August 2003 by T. Mione, plant grown in Connecticut by T. Mione. Seeds generously provided by Robert Bye 14243 grown as Mione 421 for study in the United States.
|
Figure 8. Fruits, Branches, Leaves: Photo by Robert Bye 14243 in Mexico. Slide, scanned by T. M., generously provided by Robert Bye. |
 |
Figure 9. Fruits fall at maturity and calyx (the green upside-down "funnels" in photo) remains green.
In side view, fruits are partially hidden by the calyx.
Photo by T. Mione, T. Davis 1180 grown in Connecticut as Mione 390 grown for study in the United States. |
JALTOMATA chihuahuensis Mione and R. Bye
Basionym: Saracha chihuahuensis Bitter, Repert. Spec. Nov. Regni Veg. 18: 108. 1922.
TYPE: Mexico. Chihuahua, Mpio. Casas Grandes: bei Colonia Garcia in der Sierra Madre, 2400 m, 18 July 1899. C.H.T. Townsend & C.M. Barber 91 (lectotype, MO; isotypes, K, NY, P; holotype designated by Bitter l.c. as "hb. Berol." presumed destroyed).
| Character |
Description |
Figures on this page |
Habit & Height |
Procumbent, perennial, hirsute herb. |
|
Branches, young |
|
|
older |
|
|
Leaves, blade texture & size |
|
|
| blade shape including margin |
|
|
| hairs |
|
|
| petiole |
|
|
Inflorescence |
|
|
peduncle |
|
|
pedicel |
|
|
Calyx when flowering, color & size |
|
|
| shape / position during anthesis |
|
|
| hairs |
|
|
| at fruit maturity |
|
|
Corolla color |
|
|
shape and size |
|
|
lobes / lobules |
|
|
|
|
|
|
no |
|
|
no |
|
Stamen length including anther |
|
|
| length stamens exserted beyond distal end of corolla (applicable if corolla is tubular or campanulate) |
|
|
| position of stamens: a) before anthers dehisce, b) after anthers dehisce |
|
|
| base expanded laterally? |
|
|
| filaments |
|
|
| anther color |
|
|
| anther size |
|
|
| anther mucronate/mucronulate |
|
|
| insertion of filament into anther |
|
|
| anthers of a flower open simultaneously? |
|
|
| pollen quantity per flower |
42,000 - 93,000 |
|
| pollen grain size |
|
|
Gynoecium, stigma |
|
|
Style |
|
|
| Ovary & Ovarian Disk |
|
|
Ovules per ovary |
54 - 84 |
|
Nectar |
|
|
| Herkogamy |
|
|
| Protogyny |
|
|
Fruit color (at maturity) and size |
|
|
Seeds per fruit |
no data |
no data |
Seed Size |
no data |
no data |
Chromosome number |
n = 12 |
no data |
Growability in Connecticut, USA |
no data |
no data |
How long does it take from flower to ripe fruit? |
no data |
no data |
Self-Compatible? |
yes, "abundantly sefl-set fruit in a pollinator-free greenhouse" |
Mione & Bye (1996, page 79) and J. kostyun personal communication (2016), using Mione 421 |
Seed Germination |
no data |
no data |
Ratio of pollen to ovules |
580 (48,750 / 84) |
n = 1 flower off of a plant (Bye & Linares 14243 = Mione 421)
grown in Connecticut, USA |
| Character |
Description of Jaltomata on this page |
Figures on this web page |
Finger hairs short; not dense. Leaves entire. Inflorescence 2 - 4 flowered. Peduncle 5 - 13 mm long. Pedicel 11 - 20 mm long. Flowering calyx 8 - 10.5 mm in diameter; lobe radius 3.7 - 5 mm; sinus radius 2 mm; finger hairs on abaxial face concentrated on the costa, about 10 per costa; the hairs not longer than 1 mm. Corolla rotate, pale-green; 5-lobed; 19 - 24 mm in diameter during hermaphroditic state. Stamens (not including expanded base) 3.9 - 4.0 mm long. Filaments, slender part pubescent on basal 1/5th - 1/6th. Filaments more or less parallel the style during the hermaphroditic state. Anthers 1.7 - 2.3 mm long. 32.5 - 37.5 µm in diameter. Style 3.7 - 5.6 mm long. Stigma exerted beyond distal end of anthers by 0.6 - 1.6 mm; diameter 0.38 - 0.64 mm; round in overhead outline. Infructescence 1 - 2 fruited; sometimes with a late developing flower or flower bud. Fruiting peduncles drooping; fruiting pedicels pendent; always longer than the peduncle to which they are attached. Fruiting calyx green; broadly infundibular, spreading over the fruit, covering it from above and from the side; no more than one fourth of the fruit shows from side-view. Mature fruits green in accessions Bye & Linares 14243 and Bye et al. 18329 in greenhouse (this study) and the field (R. Bye, pers. comm.). Mature fruits purple in the accession Davis 1180 in greenhouse (this study); if greenish when falls becomes purple/brown within a day or two after falling from the parent plant. Fruits always falling free from parent plant without calyx/pedicel as soon as fruit is mature.
Description based on living plants of Davis 1180 (TM 390), Bye & Linares 14243 (TM 421) and Bye et al. 18329 (TM 567).
Diagnostic Features: The filaments of all other species of section Jaltomata that I have grown for study markedly angle outwardly (inwardly on some accessions of J. repandidentata) during the first day of the hermaphroditic state while those of J. chihuahuensis only sometimes angle out, never more than 15 degrees. Jaltomata chihuahuensis is similar to J. procumbens but the former has a 5-lobed corolla and calyx mostly covering the fruit from side view, while J. procumbens has 5 corolla lobes alternating with 5 corolla lobules, and fruits fully visible from side-view in Mexico.
Distribution and Habitat: Mexico, Chihuahua, pine duff on top of white volcanic rock outcrops in shade of pines, roadside.
Uses: sweet to eat after falling from plant, sweeter than the purple form "rurusí" (Bye & Linares 14243; Bye et al. 18329). Davis and Bye (1982, p. 232) seem to have been referring to this species when they wrote, "The term metárusi is applied to the prostrate form which has lighter colored fruits and which is said to produce an edible camote from the rootstock. Metárusi is considered to be a class of rurusí. It is known to occur in a few cultivated fields and is not as common as rurusí."
Local Name: Tarahumara name is "metarsi" (Bye et al. 18329); "metárusi" (see Uses above, plant/specimen to which this name was applied not seen). The fact that this taxon is recognized by a distinct name by the Tarahumara Indians is congruent with my decision to treat it as a distinct species (see Briggs and Walters, 1984, p. 255).
Selected Specimens Examined. MEXICO. CHIHUAHUA: Rd. to La Bufa. 8.9 miles from La Bufa-Guachochi junction. T. Davis IV 1180 (CONN); mpio. Urique. al lado S de Barranca del Cobre, ca. 1 km al S de puente de Rio Urique. ca. km 61 de camino Creel-Guachoci, R. Bye & E. Linares 14243 (COLO, CONN)(TM 421).
Ethnoecology: There are two races of J. chihuahuensis, one with purple
fruits and the other with green fruits. Without mature fruits the two races
appear to be morphologically indistinguishable. Based on greenhouse and garden
studies of three accessions, we are certain that plants do not produce both
types of fruits, nor do the mature green fruits become purple with age or
exposure to sunlight. Information obtained by R.Bye from the Tarahumara during
ethnoecological studies in Chihuahua, Mexico, agrees with these observations.
Based on greenhouse studies, fruits of the purple race are typically green when
they drop and then become purple within a few days.
The two races may occupy different niches. Preliminary
observations by R. Bye are that the green-fruited race tends to occur among
white volcanic rocks in soil with higher apparent organic matter. The
purple-fruited race, however, is not found among white volcanic rocks nor in
soil with high organic matter. The rootstock ("camote") of the
purple_fruited race is eaten by the Tarahumara (Davis & Bye, 1982: 232).
Further study is needed to explore the possibility of niche specificity of the
two races.
Jaltomata procumbens (Cav.) J. L. Gentry, a
widespread species that is common within the smaller range of J.
chihuahuensis, is generally called "rurus," "rurus,"
"rurusi" or "turus" by the Tarahumara (e.g., Bye 9875 9889
COLO, Davis 1124 1127 MO; Brambila, 1976; Pennington, 1963). Jaltomata chihuahuensis,
on the other hand, is often referred to as "metrusi" or "me'tresi"
(which is a "jaltomate rastrero" or creeping jaltomate, Brambila,
1976); these names are probably derived from the Tarahumara verb 'me'trema' ('to
be creeping'). Some Tarahumara believe "metrusi" to be a class of
"rurus" (Bye, unpublished). Thus the Tarahumara recognize the
distinction between J. procumbens and J. chihuahuensis.
In most Jaltomata species, at maturity fruits
remain attached to the parent plant for at least a few days, and the accrescent
calyx darkens when the fruit ripens. The color of the calyx (purple to brown)
stands in contrast with the foliage and presumably serves, along with the
brightly colored fruit, as a beacon to diurnal fruit consumers/dispersers.
However, the fruits of J. chihuahuensis fall at maturity and the calyx
remains green. Unlike most species of Jaltomata, which exhibit
articulation at the base of the pedicel, articulation is at the attachment of
the fruit; the calyx-pedicel unit remains attached to the plant for at least
several days after the fruit drops. Human selection may result in a change from
fruits falling free of the parent plant in wild species to fruits remaining
attached to the parent plant at maturity (e.g., the nonshattering rachis of
domesticated cereals, Heiser, 1990). The consumption of fruits by the indigenous
people, perhaps subjecting the plants to human selection, has not had this
effect on J. chihuahuensis.
Fruits falling free at maturity is among the features of
the syndrome of saurochory (reptile dispersal) (Van der Pijl, 1969), and
saurochory is thus a possibility for J. chihuahuensis, if not now then
possibly prior to extensive utilization by humans. Alternatively, fruits falling
free at maturity may indicate no adaptation for dispersal, and may merely have
become fixed by genetic drift at some time when population size was small. In
any case, fruits falling free at maturity suggests that this species is not
regularly bird dispersed, although it is possible that birds feed on the fruits
that have fallen to the ground. The Tarahumara Indians consume the fruits after
they fall to the ground, and it is likely that seeds survive the human digestive
tract and so become dispersed by humans (Davis & Bye, 1982; Davis,
1986).
Humans have selected for light color in diverse
domesticates (e.g., white lupines, quinua, sheep and camels)(Heiser, personal
communication; Zeuner, 1963). It is thus possible that green-fruited mutant(s)
within J. chihuahuensis were selected by humans, giving rise to the
green-fruited race. Evidence that may be considered as strengthening this
scenario is that to humans green fruits are sweeter than purple fruits (Bye,
unpublished). Alternatively, humans may have had nothing to do with the
establishment of the green-fruited race but may merely have begun to regularly
consume the fruits.
Phylogenetic Placement: Based on chloroplast DNA
restriction sites there are two principal, sister phylogenetic groups within Jaltomata
(Mione et al., 1994). The "Mesoamerican" group, having its center of
diversity in Mexico, is widely distributed from southwestern USA to Bolivia. The
"South American" group is distributed in Andean South America, the
Greater Antilles, and the Galpagos Islands. Within the Mesoamerican clade the
rarest and most morphologically distinct species, J. grandiflora, forms the most basal
branch, and J. chihuahuensis forms the second to most basal branch (Mione
et al., 1994).
Artificial Hybridizations: Crosses were made in a
pollinator free greenhouse. Flowers used as pollen recipients were emasculated
prior to anther dehiscence. No fruits were set from 18 crosses of J. procumbens
with J. chihuahuensis (pollen source), nor from 12 of the reciprocal
cross. Two accessions of J. chihuahuensis were used (Bye 14243 green
fruits, Davis 1180 purple fruits). Six accessions of J. procumbens
from Mexico were used (Davis 1189A 1191 1124, MO; Bye 9889 10033 10084, COLO),
the latter four from Chihuahua. The following observations may be considered
controls for the above crosses. Both J. chihuahuensis and J.
procumbens abundantly self-set fruit in a pollinator free greenhouse. Within
J. chihuahuensis an uncounted number of interaccession crosses have been
successful (for study of the genetic control of fruit color), and within J.
procumbens intraccession and interaccession crosses virtually always result
in fruit set.
Taxonomy: Jaltomata chihuahuensis has been
considered to be a synonym of J. procumbens (Morton, 1938; Davis, 1986,
discussed as the prostrate morph/form of J. procumbens; Nee, 1986). The
decision to recognize this species and make the following new combination was
based on morphological characters of both living plants (in the greenhouse and
garden as well as in the field) and herbarium specimens, artificial
hybridizations and chloroplast DNA (Mione et al., 1994). Chromosome counts of n
= 12 were obtained for both J. chihuahuensis (Davis 1180) and J.
procumbens (Bye 10033). Meiocytes were stained by crushing immature anthers
with a fine dissecting needle in a drop of filtered acetic carmine.
On pressed specimens of Jaltomata chihuahuensis the
fruit is usually partially hidden by the calyx, because on living plants fruits
are pendent and the calyx (an upside-down, five-lobed funnel as in photos above) hides the fruit
from side view. On pressed specimens of J. procumbens, however, the calyx
is more likely to lie flat against the sheet because on living plants the
fruiting calyx is rotate. Based on hundreds of measurements
made on plants grown during several years, these two species cannot be
distinguished (due to overlapping ranges) with any one of the following
characters: number of flowers per inflorescence, peduncle length, pedicel
length, calyx diameter, corolla diameter, stamen length, anther length, style
width at midlength, fruit length or width, nor seed length.
Acknowledgments. I thank Tilton Davis IV for the generous
donation of his seed collection, maps and literature, the Tarahumara people for
sharing their knowledge with R. Bye. Supported
in part by an NSF grant to the University of Connecticut (T. M. and Gregory J.
Anderson).
 |
 |
 |
Figure 10, a lower left, b upper left, and c upper right. These three pictures are photos of the type specimen at P, photographed by Mione in April of 2004.
|
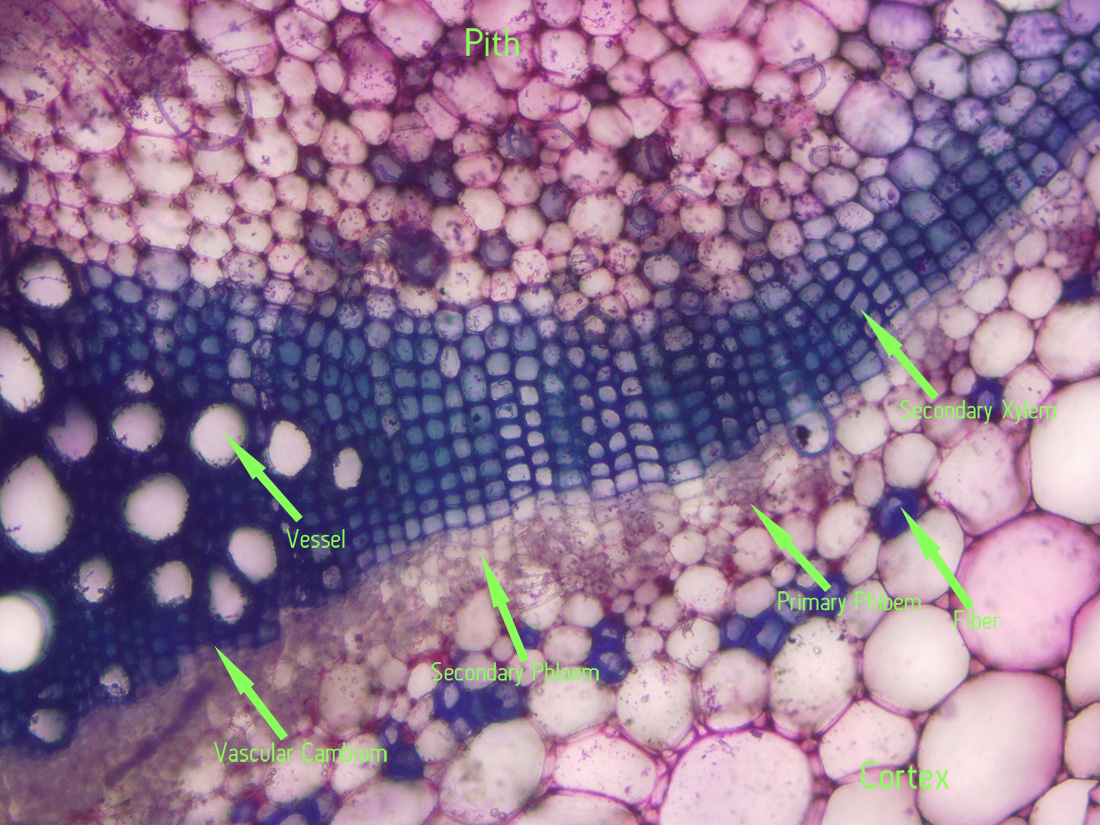 |
| Figure 11. Jaltomata chihuahensis. Cross section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. |
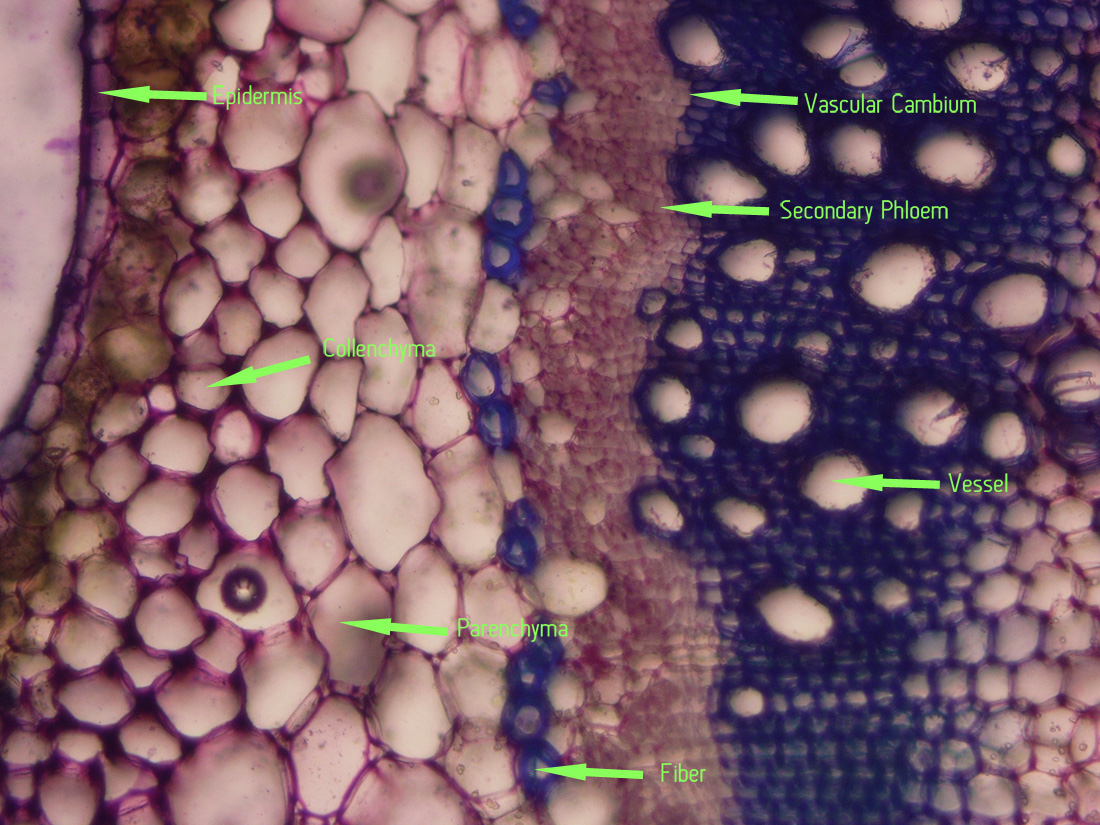 |
| Jaltomata chihuahensis. Cross section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. |
 |
| Jaltomata chihuahensis. Cross section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. |
 |
| Jaltomata chihuahensis. Cross section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. |
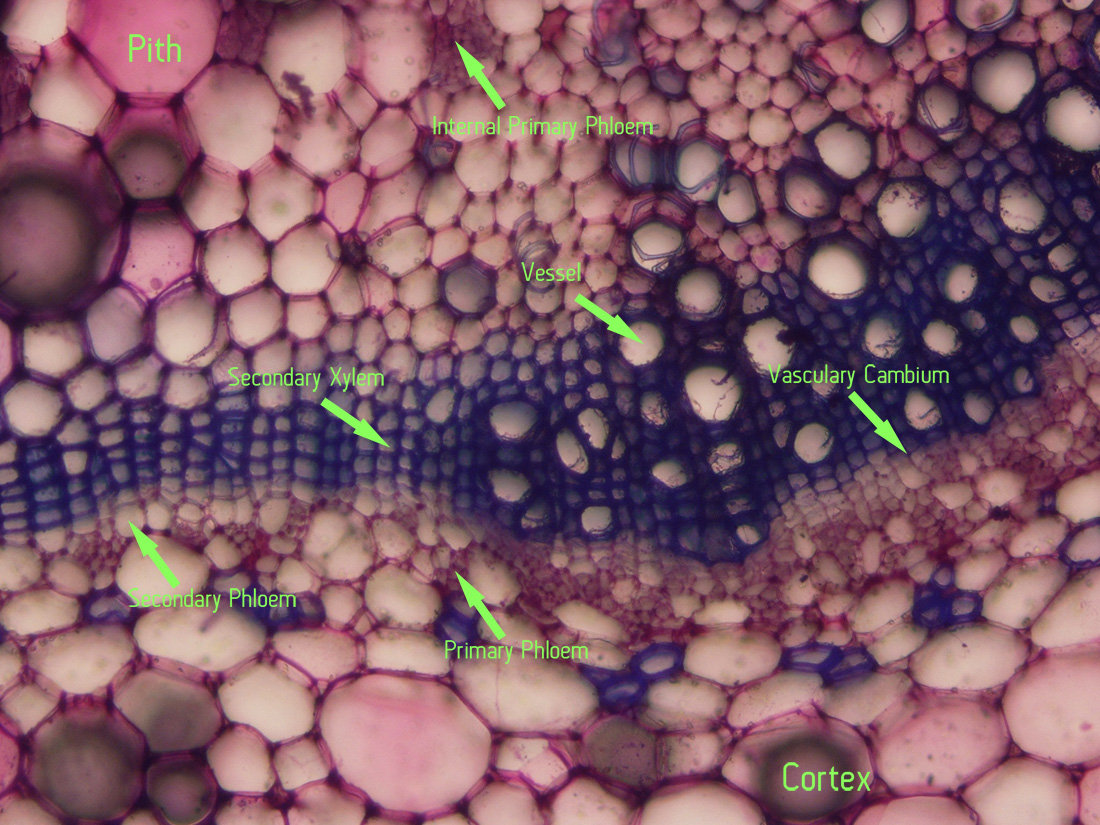
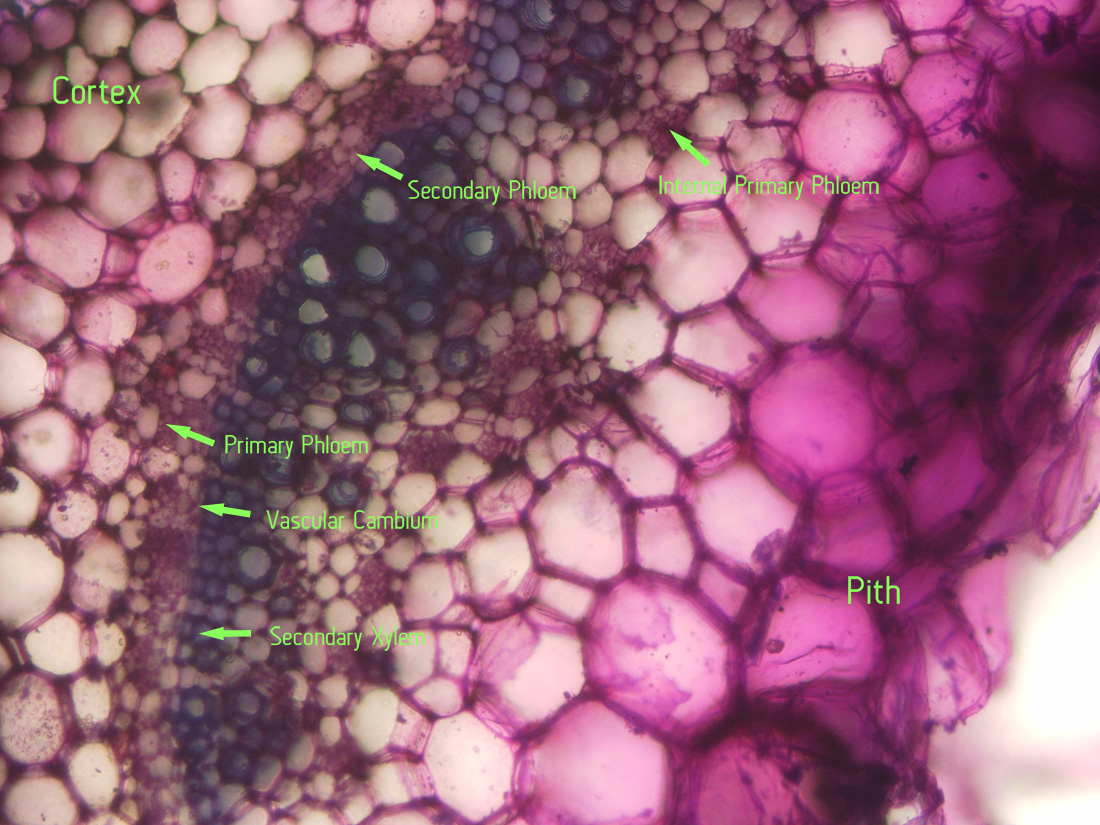 |
| Jaltomata chihuahensis. Cross section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. Note the internal primary phloem. |
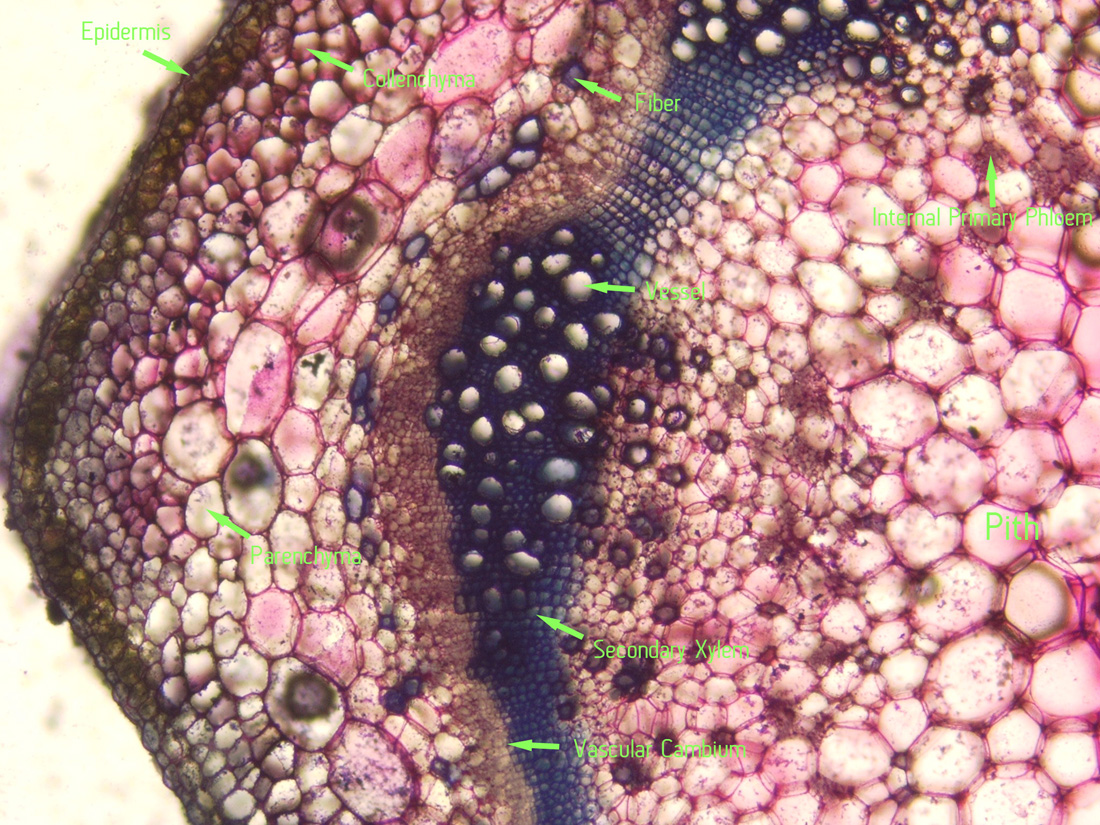 |
| Section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. Note the internal primary phloem. |
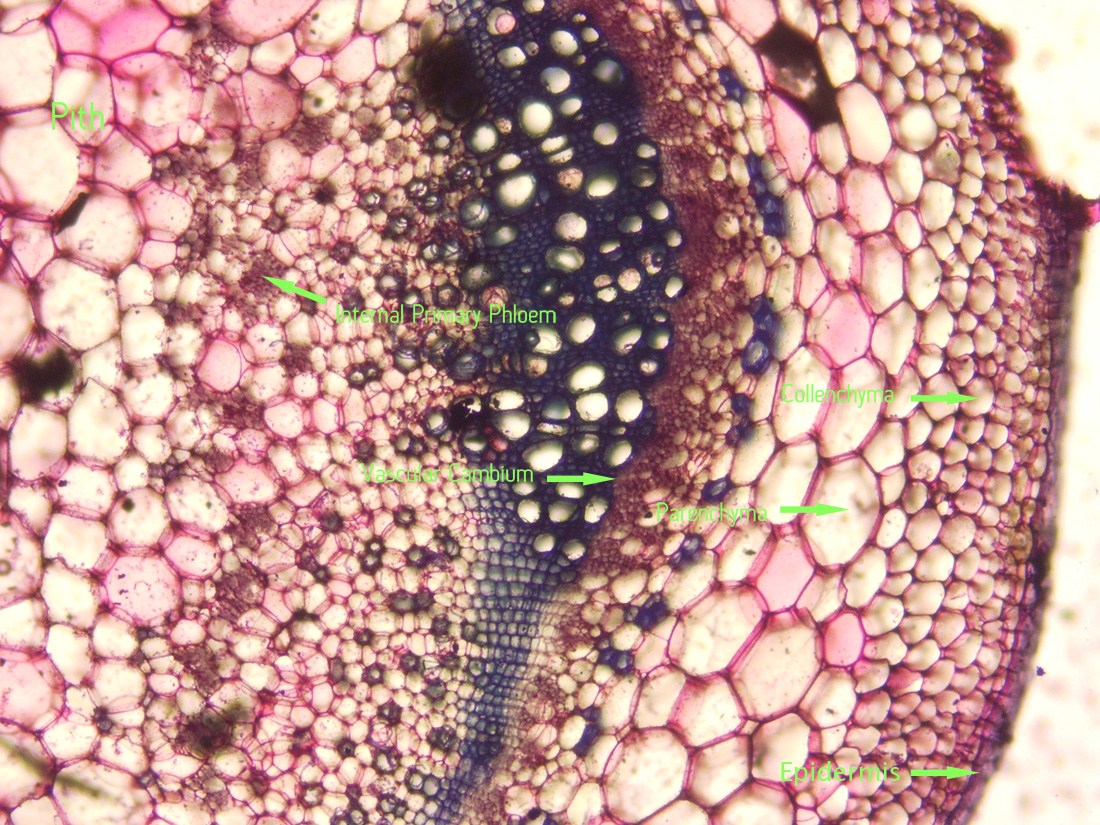 |
| Jaltomata chihuahensis. Cross section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. Note the internal primary phloem. |
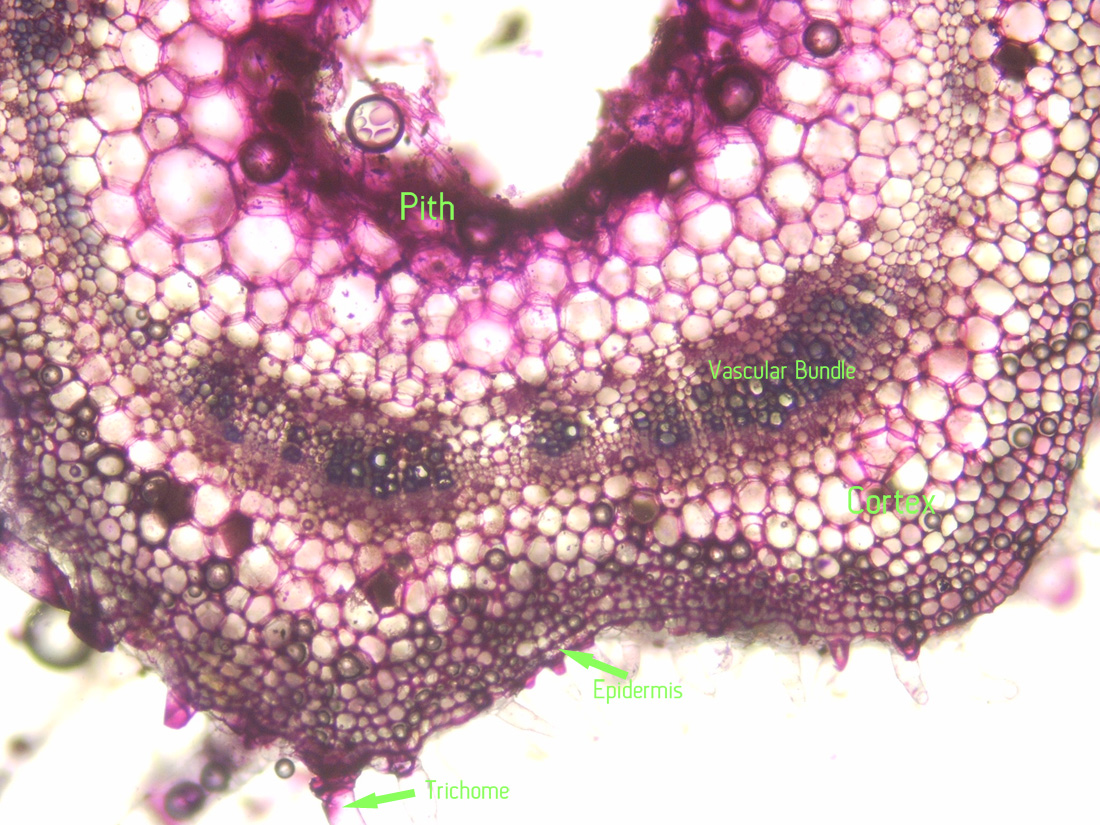
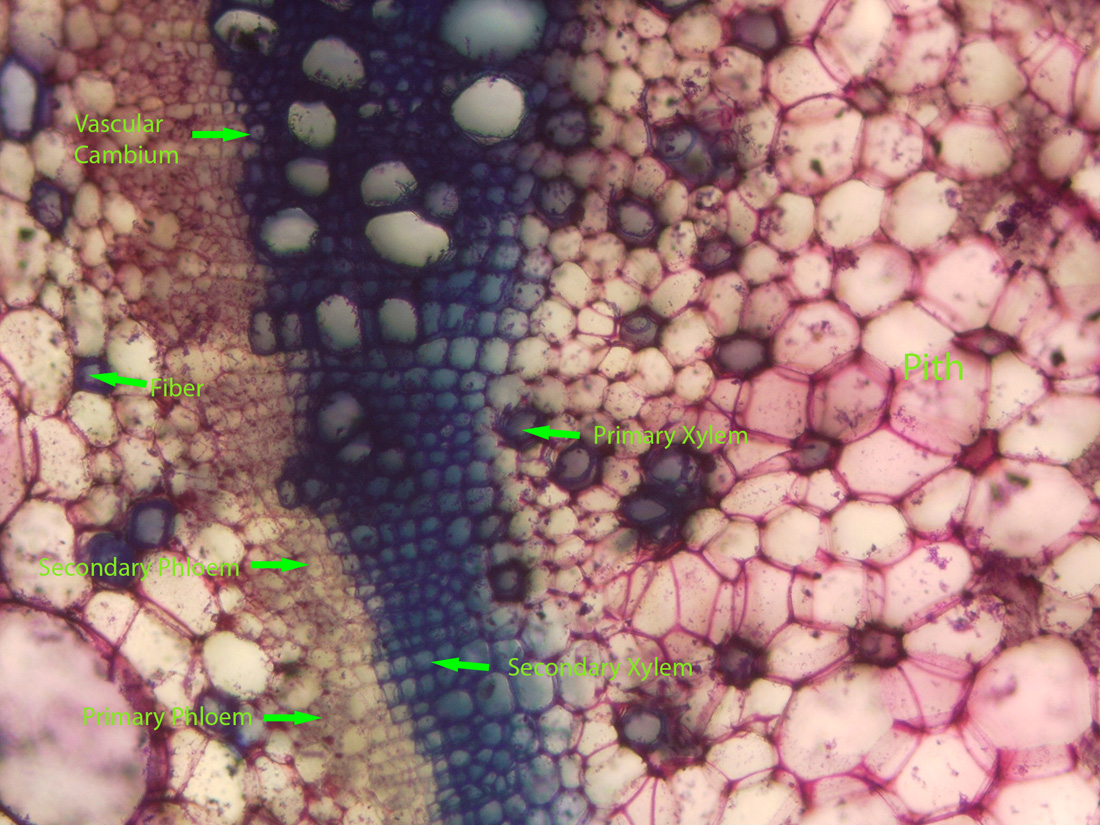 |
| Jaltomata chihuahensis. Cross section (by hand with a razor blade, stained with toluidine blue) and photography by Christopher L. Chan; Mione 390 grown for study in the United States. |
Literature Cited
Brambila, D. 1976. Diccionario raramuri-castellano (tarahumar). La Obra
Nacional de la Buena Prensa, Mxico, D.F.
D'Arcy, W. G., T. Mione & T. Davis IV. 1992. Jaltomata grandiflora
(Solanaceae): a rare Mexican species. Novon 2: 190-192.
Davis, T. 1986. Jaltomata in the Tarahumara Indian region of northern
Mexico. Pp. 405-411 in W. G. D'Arcy (editor), Solanaceae Biology and Systematics.
Columbia University Press, New York.
_____ & R. A. Bye, Jr. 1982. Ethnobotany and progressive domestication of
Jaltomata (Solanaceae) in Mexico and Central America. Econ. Bot. 36: 225-241.
Heiser, C. B. 1990. Seed to civilization: the story of food. Harvard Univ.
Press, Cambridge.
Knapp, S., T. Mione & A. Sagstegui A. 1991. A new species of Jaltomata (Solanaceae)
from northwestern Peru. Brittonia 43: 181-184.
Mione, T. 1992. Systematics and evolution of Jaltomata (Solanaceae).
Ph.D. dissertation, University of Connecticut Storrs, CT.
_____, G. J. Anderson & M. Nee. 1993. Jaltomata I: circumscription,
description and new combinations for five South American species. Brittonia 45:
138-145.
_____, & F. G. Coe. 1992. Two new combinations in Peruvian Jaltomata
(Solanaceae). Novon 2: 383-384.
_____, R. G. Olmstead, R. K. Jansen & G. J. Anderson. 1994. Systematic
implications of chloroplast DNA variation in Jaltomata and selected
physaloid genera (Solanaceae). Amer. J. Bot. 81: 912-_918.
Morton, C. V. 1938. Notes on the genus Saracha. Proc. Biol. Soc. Wash.
51: 75-_77.
Nee, M. 1986. Solanaceae I. In Flora de Veracruz, fascculo 49. Instituto
Nacional de Investigaciones sobre Recursos Biticos, Xalapa, Veracruz, Mxico.
Pennington, C. W. 1963. The Tarahumar of Mexico: Their environment and
material culture. Univ. of Utah Press, Salt Lake City.
Van der Pijl, L. 1969. Principles of Dispersal of Higher Plants.
Springer_Verlag, Berlin.
Zeuner, F. E. 1963. A history of domesticated animals. Harper & Row, New
York.