 |
Figure 1. Branches, inflorescences and leaves. The corolla is pale-green prior to anthesis (to the right of center) but after it opens it becomes white and remains white. In the flower at the center all anthers have dehisced while in the flower at right some of the anthers have dehisced while others have not. The stigma is green and the style is whitish; the upper half of the ovary is green, and the lower half of the ovary is surrounded by an orange disk, presumably the source of nectar.
This plant was grown from seed of the type collection by Thomas Mione in Connecticut, USA; photo by Thomas Mione. |
 |
Figure 2. Flower at left was pressed in Ecuador (type collection) and the flower at right is fresh (not pressed) and was removed from a plant grown from seed of the type collection. The size difference we see between these flowers is almost certainly due to growing conditions, not genetic differences. The plant in Ecuador (at left) was probably competing for nutrients, water and sun while the plant at right was not competing. The units along the left edge are mm. Photo by Thomas Mione in the laboratory of Gregory J. Anderson. |
 |
| Figure 3. Ripe fruits, leaves, stems (Mione et al. 710, photo by Thomas Mione, original print was scanned for this page). |
 |
| Figure 4. The white ruler (21 cm long) is resting on Jaltomata lojae and the leaves at the bottom middle of the photo are also J. lojae. Peru, Department Piura, province Huancabamba (Mione et al. 811, photo by Thomas Mione). |
| Character |
Description of Jaltomata lojae |
Figures on this web page |
Habit & Height |
Perennial shrub |
|
Branches, young |
branches and leaves densely villous, bearing finger hairs, these sometimes gland-tipped, 0.15 - 6 mm long, becoming less villous with age |
|
older |
to 14 mm diam (Mione et al. 813) |
|
| Leaves, size |
Leaves alternate, often geminate, to 15 cm long X 6 cm wide |
|
| shape |
ovate, the margin entire to sinuate-dentate, undulate |
|
| hairs |
branches and leaves densely villous, bearing finger hairs, these sometimes gland-tipped, 0.15 - 6 mm long, becoming less villous with age |
|
| petiole |
to 4.5 cm long |
|
Inflorescence |
umbelliform, to 9-flowered |
|
peduncle |
5 - 9 mm long (at flowering) having a dense covering of erect finger hairs to 1.5 mm long, some hairs gland-tipped |
|
pedicel |
8 - 11 mm long (at flowering) having a dense covering of erect finger hairs to 1.5 mm long, some hairs gland-tipped |
|
| Calyx at flowering |
Calyx light green, 9.5 mm in diameter, strongly reflexed, abaxially villous with gland-tipped finger hairs, forked hairs, and dendritic hairs all 1 - 2 mm long, and abaxially and adaxially with stalked multicellular glands 62 - 80 µm long |
|
| at fruit maturity |
at fruit maturity calyx diameter to 12 mm. |
|
Corolla color |
white with two green, proximally positioned maculae straddling the radial vein to each corolla lobe |
|
shape and size |
infundibular when partially open, crateriform-rotate when fully open, 25 - 27 mm in diameter on plants I grew, 18 mm in diameter on the holotype measured after pressed/dried |
|
lobes/lobules |
5 prominent lobes alternating with 5 small lobules |
|
hairs |
adaxially glabrous, abaxially with sparsely but evenly distributed stalked multicellular glands 62 - 80 µm long. Two classes of hairs extend out from the corolla margin: marginal hairs to 110 µm long, and submarginal (attached abaxially) hairs to 0.5 mm long. |
|
|
no |
|
Stamen length |
5 mm long |
|
| filaments |
filaments on living plants angling away from style, and slightly excurved, villous on basal 1/4 of the length with unpigmented finger hairs to 1.2 mm long |
|
| anther color |
|
|
| anther size |
anthers 2.1 - 2.5 mm long prior to dehiscing, 1.6 - 1.8 mm long after dehiscing, when corolla is fully open exserted out of corolla 2 mm, otherwise included |
|
| anther mucronate |
|
|
| anthers of a flower open simultaneously? |
The anthers of a flower either dehisce a few at a time over the course of several hours, or two or three of the anthers dehisce one day and the others dehisce the next day |
|
| pollen quantity |
|
|
| pollen grain size |
see Table 3 on this page
|
|
| corona |
no |
|
Stigma |
shallowly bilobed, 0.4 to 0.75 mm wide on pressed specimens, broader than the style, the papillae 30 - 60 µm long |
|
Style |
9 - 11 mm long, slender and straight, exserted 3 - 8 mm beyond the anthers |
|
Ovary |
nectar disk orange |
|
Ovules per ovary |
|
|
| Nectar |
transparent |
|
| Herkogamy |
yes, the stigma is located 3 to 8 mm from the anthers |
|
| Protogyny |
yes |
|
Fruit color (at maturity) and size |
orange |
|
Seeds per fruit |
numerous |
|
Seed Size |
ovate to reniform, 1.3 - 1.6 mm long |
|
Chromosome number |
n = 12 (nine counts) |
Chromosome counts were made with meiocytes of immature anthers stained with acetic carmine. |
Growability in Connecticut, USA |
was easy in greenhouse |
|
How long does it take from flower to ripe fruit? |
no data |
|
Self-Compatible? |
|
|
Seed Germination |
|
|
| Character |
Description of Jaltomata lojae |
Figures on this web page |
Specimens of Jaltomata lojae examined
(grouped first by country then chronologically ordered)
| Country |
Province |
|
Locality |
elevation m |
habitat |
date |
collector |
Data Entry |
| Ecuador |
Chimborazo |
|
Cañon on the río Chanchan, directly above the village of Huigra |
|
open deforested slope with small patches of scrub in the draws |
|
Camp E-3498 (NY) |
Aug 2011 |
| Ecuador |
Loja |
|
Lugma Huycu 12 km north of Saraguro,
79 15' W, 03 34' S |
2,100 |
no data |
19 Jan 1989 |
Ellemann 66799 (NY) |
Aug 2011 |
| Ecuador |
Loja |
|
Cerro Sozoranga, Colaisaca-Utuana, km 0.5, 4 19' 14" S, 79 41' 16" W |
|
hedges |
24 Apr 1994 |
Jørgensen et al. 567 (NY) |
Aug 2011 |
| Ecuador |
Loja |
|
Sevillán, 3 32' 10" S
79 22' 30" W |
2,740 |
potrero |
26 Mar 1995 |
V. Van den Eynden & E. Cueva 342 (NY) |
Aug 2011 |
| |
Department |
Province |
|
|
|
|
|
|
| Peru |
Piura |
Huancabamba
|
Abra de Porculla, entre Olmos y Jaén |
|
ladera con monte bajo |
|
R. Ferreyra 15667 (K, US) |
Aug 2011 |
| Peru |
Piura |
Huancabamba |
46-48 km E of Olmos on road to Pucara, just W of top of Abra de Porculla |
|
roadside, lower montane dry forest |
|
A. Gentry, M. Dillon, et al. 22684 (NY) |
Aug 2011 |
| Peru |
Piura |
Huancabamba |
Perculla |
2,200 |
zonas húmedas y arcillosos |
2 May 1981 |
S. Llatas & J. Laes 631 (HUT not seen, MO, NY) |
Aug 2011 |
| Peru |
Piura |
Huancabamba |
Carretera entre Canchaque y Huancabamba, km del 16 al 25 desde Canchaque |
|
no data |
|
C. Díaz S. & S. Baldeón 2395 (MO, NY) |
Aug 2011 |
| Peru |
Piura |
Huancabamba |
Highway Olmos to Chamaya |
|
quebrada |
|
S. Leiva G., T. Mione, L. Yacher 3339;
Mione, Leiva & Yacher et al. 710 |
March 2009 |
| Peru |
Piura |
Huancabamba |
road to Huancabamba, 5 22' 00" S, 79, 33' 52" W |
|
roadside |
|
S. Leiva G., T. Mione, L. Yacher 5104;
Mione, Leiva & Yacher et al. 811 |
Aug 2011 |
| Peru |
Piura |
Huancabamba |
5 21' 26.7 to 29" S,
79, 34' 31 to 31.6" W |
1,986 |
roadside and trailside |
|
S. Leiva G., T. Mione, L. Yacher 5106;
Mione, Leiva & Yacher et al. 813 |
Aug 2011 |
 |
| Figure 5. Corolla maculae (green) are visible through the corolla. Mione et al. 710, photo by Thomas Mione |
 |
| Figure 6. Style is exserted. Plant grown from seed of the type collection by and photo by Thomas Mione. |
 |
Figure 7. Comparison of the type collection (right) and a plant grown in Connecticut from seed of the type collection (left). The plant at left was pressed in Connecticut, after being grown from seed of the type collection; the plant at right is the type collection and was pressed in Ecuador. The plant at right was probably competing for nutrients, water and sun while the plant at left was not competing. Plant at left grown by and photo by Thomas Mione in the laboratory of Gregory J. Anderson. |
 |
| Figure 8. Flower. Note curved filaments, and that anthers have dehisced. Stamens and corolla tissue were removed to allow view of the ovary including the orange disk that surrounds the base of the ovary. The orange disk around the base of the ovary is probably the nectary. The style disappears distally because of poor lighting, but the stigma (green) is showing at the top of the photo. The lilac-colored pigmentation immediately distal to the corolla maculae (green) was not present in the type specimen. Smallest units are mm. Photo by Thomas Mione, Mione et al. 813. |
 |
| Figure 9. Leaves. Upper side showing of larger leaf on left and the two leaves on the right. Numbered units are cm. Photo by Thomas Mione, Mione et al. 813. |
 |
| Figure 10. Leaves. Upper side showing of larger leaf on left and the two leaves on the right. Numbered units are cm. Photo by Thomas Mione, Mione et al. 813. |
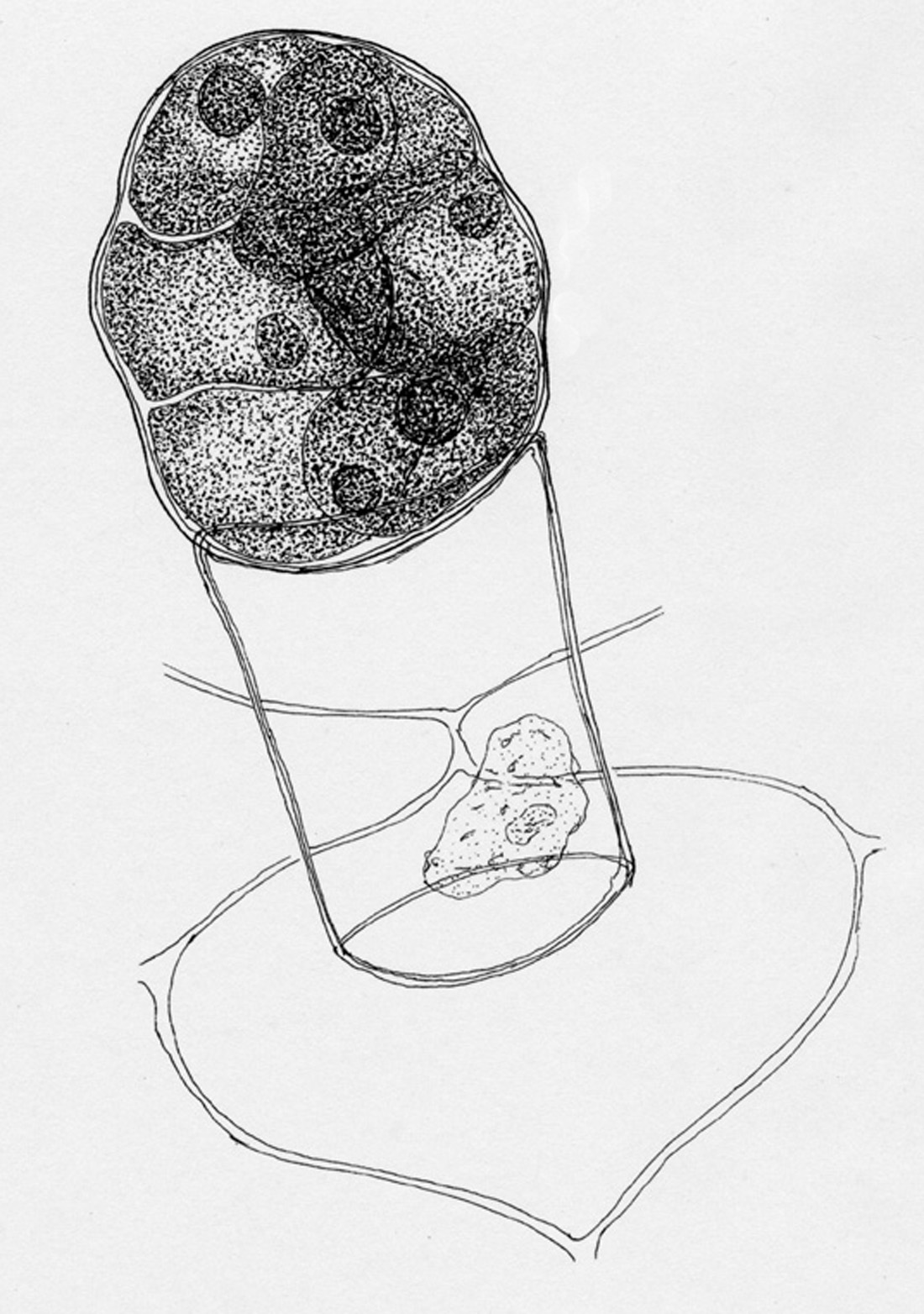 |
| Figure 11. Multicellular gland, about 1/10th of a mm long, located on the abaxial face of the corolla and both faces of the calyx. The multicellular head stains darkly with neutral red while the stalk cell does not. Illustration by Luis A. Serazo. |
 |
| Figure 12. Mione et al. 811 |
 |
| Figure 13. Mione et al. 811 |
 |
| Figure 14. Mione et al. 811 |
I thank David M. Spooner for seeds of the type collection, and Janet R. Sullivan for review of the manscript that became the protologue.