The combination of large somewhat coriaceous leaves, and flowers that lack protogyny, last only a single day and have curved styles, contributed to the recognition of this taxon as a distinct species.
In this description trichomes are not gland-tipped unless indicated as such. Finger hairs are uniseriate, unbranched and multicellular. Branchlet hairs have multiple termini (Seithe, 1979). Gland-tipped finger hairs (on the Jaltomata of Costa Rica found only on the adaxial face of the corolla) have an expanded terminal cell that stains densely with neutral red. As well, the multicellular head of the stalked glands (common on the abaxial faces of the perianth) also stains densely with neutral red. Pollen grain diameter was measured after staining pollen 30 minutes in "cotton blue" stain; anthers were stored in 70% ethanol prior to staining. Floral phenology was observed in the field, and on healthy plants grown for study both outdoors, and indoors under lights. Pollen grains were counted using the method of Anderson and Symon (1989).
| Habit & Height |
Erect glabrous herb to 1.17 m high x 0.91 m across. |
|
| Stems |
Stems hollow, green to purple, strongly angulate with 4 to 5 raised longitudinal ridges, to 3.5 cm diameter at or just above base. |
|
| Older Branches different from younger? |
|
|
| Leaf Size |
alternate, often geminate, to 30 cm long, , the margin entire or nearly so and on smaller, younger leaves sparsely ciliolate with finger hairs to 0.1 mm long; petiole to 2.5 cm long. |
|
| Leaf Shape |
the blade elliptic, the proximal most 13—16% of the blade cuneate, the tip usually acuminate, the length to width ratio 1.5 to 1.8, the margin entire or nearly so |
|
| Leaf texture and hairs |
the faces glabrous and lustrous, somewhat coriaceous, younger leaves sparsely ciliolate with finger hairs to 0.1 mm long |
|
| Inflorescence |
axillary or arising from a stem dichotomy, umbellate, to 41-flowered (including very small buds) on plants cultivated for research |
|
| Peduncle & Pedicel |
Peduncle to 2.1 cm long and pedicel to 2.3 cm long; both green, glabrous, and having raised longitudinal ridges. |
|
| Calyx at Flowering |
green, 5.2—7.4 mm from pedicel to tip of lobe, 12--14 mm in diameter, the midrib raised, ciliolate near apex with 1 to 3 celled finger hairs 38—138 µm long, the hairs usually simple but occasionally with more than one terminus, abaxially with abundant stalked glands 60—75 µm long. |
|
| Corolla shape and size |
rotate, 17—20 mm (-25 mm, cultivated) in diameter |
1 |
| Corolla lobes/lobules |
5-lobed, |
1 |
| Corolla color and hairs |
light-green with dark-green maculae collectively stellate in outline, the margin ciliolate with branchlet hairs to 190 µm long, adaxially densely pilosulose with erect, gland-tipped (droplet-tipped on fresh flowers) finger hairs, (4) 5—6 celled and 210—290 µm long, abaxially with abundant stalked glands 70—80 µm long. |
1 |
| Stamens |
4.5—5 mm long; filaments straight or nearly so, nearly glabrous with unpigmented finger hairs to 0.3 mm long only on expanded base |
1 |
| Anther length & color |
anthers1.4—1.9 x 1.0—1.8 mm, yellow |
|
| anthers of a flower open simultaneously? |
yes |
|
| Pollen Grains |
25—27.5 µm in diameter (n = 59 grains);
74,000—149,000 grains per androecium (n = 2 androecia)
42,500 grains per androecium (n = 1 androecium, 2011) |
The 2011 pollen count was done by CCSU student Elisabeth dos Santos |
| Stigma |
green, capitate,with a shallow medial groove (when living), 0.33—0.51 mm wide (when pressed), the papillae to 28 µm long (living) |
|
| Disk around ovary |
off-white to light orange disk girdling the base of the dark green ovary |
|
| Style length |
style 4.2—5.1 mm long, curving and therefore orienting the stigma to the side (see photos above) |
|
| Ovules per ovary |
158—168 per ovary (n = 2 ovaries) |
The ovule count of one locule was doubled to obtain the total ovules per ovary. |
| Nectar |
Translucent nectar droplets form where the corolla and androecium meet. This nectar is likely produced by the ovarian disk and then, taking the path of least resistance, seeps through the bases of the stamens on to the corolla. |
|
| Herkogamy? |
Yes, the dehsiced anthers and stigma are about 4 mm away from each other during the first and only day the flower is open |
|
| Protogyny |
No |
|
| Fruits |
dark black-blue at maturity, subspherical, to 14 mm in diameter, remaining attached to parent plant (not dropping at maturity). |
|
| Calyx at fruit maturity |
to 26 mm in diameter, stellate, green, rotate to slightly reflexed, not hiding fruit in side view. |
|
| Infructescence |
|
|
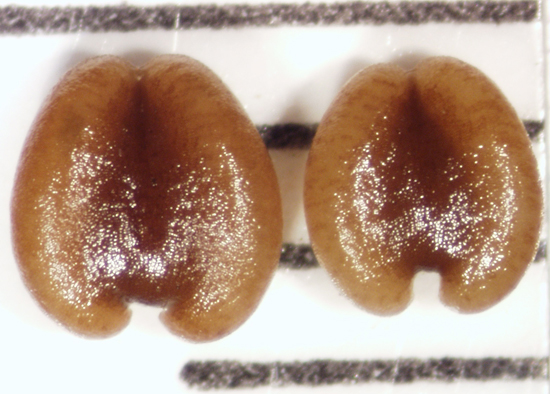
| Seeds |
subovate, alveolate,1.8—1.98 X 1.41—1.62 X 0.45—0.54 mm thick,
to 146 per fruit. |
|
| Seedlings |
have gland-tipped stem hairs (2009).
2010: All hairs uniseriate unpigmented. I don't see droplets of fluid or viscous secretion at the tips of the hairs (dissecting microscope), but I do see that some seedling hairs have a terminal, bulbous cell that has a nucleus and other hairs taper to a point (compound microscope, 200X) |
|
| Chromosome number |
2n = 24. Chromosomes were counted with the method of Bernardello & Anderson (1990); to obtain root tips seeds were sown on wet filter paper in a petri dish at room temperature under ambient light. |
|
| Growability in Connecticut, USA, outdoors |
Easy |
|
Collections of Jaltomata darcyana including Geographic Distribution and Altitudes (chronological order)
| Country |
Primary Political Region |
Locality |
elevation
m |
habitat |
date |
collector |
data entry |
| Costa Rica |
Guanacaste |
Santa Rosa |
no data |
in shade |
Sept 1971 |
Callaway 294 (US) |
August 2010 |
| Costa Rica |
Puntarenas |
TYPE SPECIMEN. Nicoya Peninsula, Curú, 90 46’ 50” N, 840 56’ 05” W, |
98 |
edge of pasture |
11 Jan. 2000 |
T. Mione & L. Yacher 694 (holotype, NY!; isotypes: CONN!, CR!, MO!) |
August 2011 |
| Nicaragua |
Esteli |
Salto de Estanzuela |
no data |
humid canyon |
13 Aug 1976 |
J. S. Hall 7685 (CR); Jamie Kostyun wrote (personal comm. 2014) "definitely looked like J. darcyana" |
July 2014 |
| Mexico |
Guerrero |
San Luis Acatlán, Yoloxóchitl |
591 |
vegetación secundaria de bosque tropical subcaducifolio |
2017 |
K. Velasco Gutiérrez 40481 (F) |
July 2023 |
| Mexico |
Guerrero |
San Luis Acatlán, Yoloxóchitl, eastern edge of town |
624 |
alongside main road |
18 Oct 2010 |
J. D. Amith et al. 1858,
grown as Mione 313 |
August 2011 |
| Mexico |
Jalisco |
19 39' N, 104 04' W. Tuxcacuesco, Centro de asistencia del LNLJ Zenzontla |
820 m |
Veg. sec. de bosque tropical subcaducifolio |
9 Aug 1988 |
L. Robles 600 (WIS) |
Apr 2014 |
| United States |
Illinois, Chicago |
Little Village neighborhood, part of South Lawndale (41° 50' N, 87° 42 W) |
|
home garden of a woman who brought the seeds with her from Michoacán at least 20 years earlier, and grows it every year |
2018 |
John Taylor (without collection number), grown by T. Mione as 311 |
Oct 2018 |
The following information will be organized into the table above
Paratypes. COSTA RICA. Guanacaste: Santa Rosa National Park, ca. 10 50’ N., 85 37’ W., 0--320 m, 16 July 1981, Janzen 12113 (MO), and 10 50’ N., 85 35’ W., 8 Nov. 1981, Janzen 12125 (F);
Nicoya Peninsula, Cantón Nandayure, Pilas de Bejuco, Quebrada Seca, 9o 52’ 01” N., 85o 20’ 45” W., 40 m, 15 Oct. 1994, Estrada & Rodríguez 242 (CR, INB).
Puntarenas: Reserva Biológica Carara Estación Quebrada Bonita, 9 46’ N., 84 36’ W., 30 m, 26 June 1990, Bello & Rojas 2295 (CR, F, INB);
Nicoya Peninsula, Curú, pastures S of entrance road, mouth of Marianas Canyon, 50 m, 13 Sep. 1995, Sanders & Baker 17892 (F, NY).
|