
Figure 1.
Three anthers have dehsiced,
two (of shorter stamens)
have not yet
dehisced.
Photo by Thomas Mione
Jaltomata biflora (R. & P.) Benítez |
Peru |
revised February 2022 |
| Link to Jaltomata homepage | The information on this page may be cited as a communication with professor Thomas Mione, Central Connecticut State University, Biology Department, Copernicus Hall, 1615 Stanley Street, New Britain, Connecticut 06050-4010, United States of America. |
Link to list of Jaltomata species having edible fruits,
including this species |
Link to local names including of this species |
Link to list of Jaltomata species having red / orange nectar |
|
Figure 1. Three anthers have dehsiced, Photo by Thomas Mione |
Figure 2. The above photo and the photo on the right |
|
 |
Figure 3. Photo of the type specimen
|
Saracha biflora Ruiz & Pav. Flora Peruviana 2: 42-43, t. 179. 1797-99 [not Atropa biflora, a synonym of J. bicolor].
| Character | Description of Jaltomata biflora | Figures on this page |
Habit & Height |
Shrub, sprawling and woody, can be seen growing down a steep slope (rooted higher than much of the plant) |
Macbride (1962 page 31, as Saracha biflora) incorrectly described this species as herbaceous. |
Branches, young |
In nature: green, terete, pubescent, the hairs finger, forked or dendritic, non-gland-tipped; Notes that must be from greenhouse: very densely hairy, gland-tipped finger hairs |
|
older |
6 - 7 mm diameter, terete | |
Leaves, size |
||
| shape | ovate, the apex acute, entire to nearly entire | |
| hairs | the blade's upper face: pubescent with non-gland-tipped hairs the blade's lower face: puberulent |
|
| petiole | ||
Inflorescence |
1 - 2 flowered (my observations) "2- or rarely 3-flowered" (Macbride 1962, page 31, as Saracha biflora) |
|
peduncle |
green, to 15 mm | |
pedicel |
green, to 15 mm | |
Calyx at flowering |
green, dense indument of gland-tipped finger hairs | |
| shape / position when flowering | rotate | 6 |
| at fruit maturity | ||
Corolla color |
green | |
shape and size |
urceolate, 1.3 cm long, the base to 1.4 cm in diameter, the limb becoming revolute | |
lobes/lobules |
10-total: narrowly triangular lobes alternating with inconspicuous lobules | |
hairs
|
dense indument of gland-tipped finger hairs | |
Stamen length including anther |
22 - 23 mm | 7 |
| length stamens exserted beyond distal end of corolla (applicable if corolla has a well-defined tube) | 7.5 - 14 mm | based on both plants I grew and photos of the type specimen at P |
| yes | 4 | |
| base expanded laterally? | yes, as seen in ventral view | 5 |
| filaments | pubescent on proximal 36 - 39 % of length | 5, 7 |
| anther color | purple before dehiscence | |
| anther size | ||
| anther mucronate / mucronulate | no | |
| insertion of filament into anther | ||
| anthers of a flower open simultaneously? | During April, two to four of the anthers of a flower dehisced one day and the others dehisced the morning of the next day (n = six flowers). In May, however, when daytime temperatures in the greenhouse were noticeably warmer, all of the anthers of a flower dehisced in one day by 10:00 am (n = four flowers). | Mione et al. 2001 |
| pollen grain size | ||
| corona | no | |
Stigma |
shallow medial groove, shallowly bilobed | |
Style |
12 -- 18 mm, with the extremes of this range present on the same plant at the same time | 4 |
Ovary |
ovarian disk | 4 |
Nectar |
Nectar was clear at first, and although inconspicuous, could be seen through the corolla wall. Nectar became more conspicuous toward the end of the hermaphroditic phase: it turned amber to orange in color, as viewed both through the corolla wall and when removed with a clear pipette tip. | Mione et al. 2001 |
| Herkogamy | yes prior to anther dehiscence | |
| Protogyny | yes | |
Fruit color (at maturity), size, and seeds per fruit |
orange 11 X 16 mm containing 119 seeds (collected in wild, Mione et al., 876) 11 X 19 containing 167 seeds (collected in wild, Mione et al. 890) |
|
Seeds per fruit |
161 and 166 |
n = 2 fruits collected in Peru, Mione et al. 2001 |
Seed Size |
||
| Character | Description of Jaltomata biflora | Figures on this page |
| Character | Description of J. biflora | |
Growability in Connecticut, USA |
Easy to grow | Mione et al. 2001 |
How long does it take from flower to ripe fruit? |
no data |
|
Flowers Closing For The Night? |
no | Mione et al. 2001 |
Self-Compatible? |
yes | Mione et al. 2001 |
Seed Germination |
After no germination for weeks while being kept moist, I let the potting mix (in a cup) containing the seeds dry out completely and then put the cup in the mist room, and then several seeds germinated | |
Pollen quantity per flower |
104,167 n = 1 flower, Elisabeth
dos Santos 2011 n=1 flower, Kenneth C. Plourd 2016 167,500 grains |
|
Ovules per ovary |
174 | the ovules of one locule were counted by T. M. and then doubled |
Ratio of pollen to ovules |
963 | 167,500 grains/174 ovules from the same flower |
Chromosome number |
no data |
|
| Character | Jaltomata biflora |
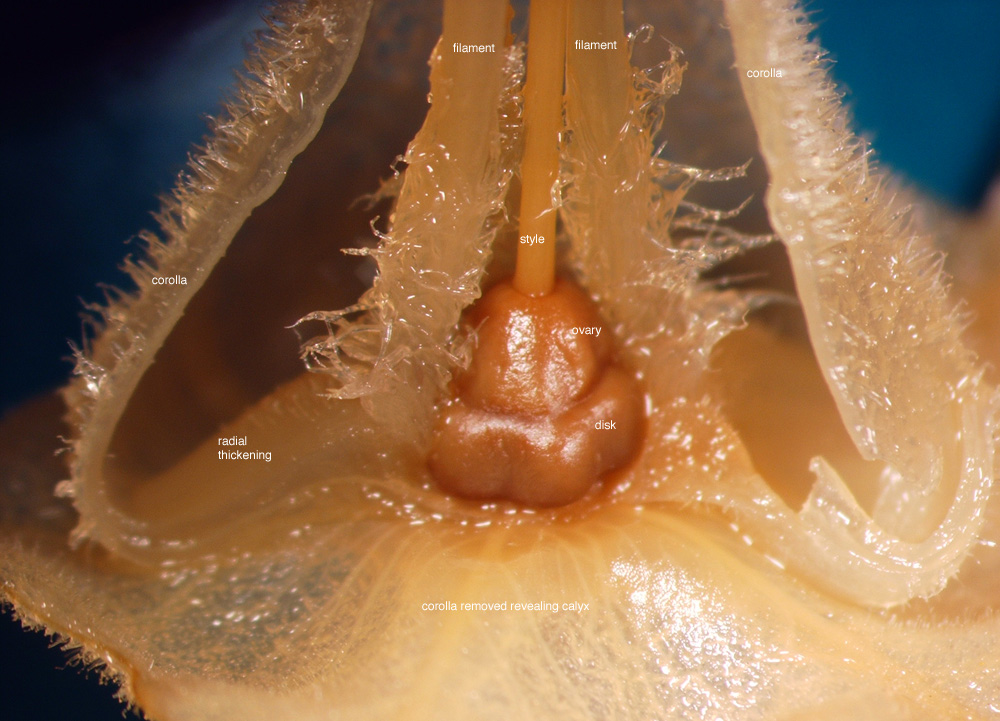 |
| Figure 4. Longitudinally dissected flower of Jaltomata biflora; dissection and photo by Melissa Luna, D. Mugaburu 5 grown as Mione 608, flower from a plant grown in Connecticut and then preserved in 70% ethanol. |
Country, Department Province |
Locality | elevation m | habitat | date | collector | Data Entry |
| Peru, Junín, Tarma | "huasa-huasi" on the label of the types and "Huassahuassi" in the protologue. Modern: Huasahuasi | Dombey | April 2009 |
|||
| Peru, Junín, Tarma | Carpapata, above Huacapistana | 2,700 - 3,200 | edge of forest | June 1929 |
E. P. Killip & A. C. Smith 24396 (NY, US) | April 2009 |
| Peru, Junín, Tarma | walked up from town of Huasahuasi | 2,965 - 3,030 | sunny, among shrubs | June 1996 |
Daniel Mugaburu 5 & 6, grown as Mione 608 | March 2009 |
| Peru, Junín, Tarma | up from town of Huasahuasi 11 14 58.9" S, 75 40' 57.4" W |
3,109 | among shrubs | 23 May 2016 | T. Mione, S. Leiva G. & L. Yacher 876 = S. Leiva G., T. Mione & L. Yacher 6071 |
July 2016 |
| Peru, Junín, Tarma | up from town of Huasahuasi 11 15 34.6" S, 75 39' 43.7" W |
2,849 | roadside nearly vertical slope | 15 March 2018 | T. Mione, S. Leiva G. & L. Yacher 889 = S. Leiva G., T. Mione & L. Yacher 6751 |
March 2018 |
| Peru, Junín, Tarma | roadside north of Jauja 11 23 20.7 S, 75 41 05.9 W |
2,865 | vertical man-made rock wall on side of highway | 15 March 2018 | T. Mione, S. Leiva G. & L. Yacher 890 = S. Leiva G., T. Mione & L. Yacher 6752 | March 2018 |
The following is modified from Mione et al. 2001: The fruits of Jaltomata biflora are at least occasionally eaten. Until this study Jaltomata biflora (R. & P.) Benítez was known almost exclusively from the type specimens collected by Dombey in the late 1700s and the inaccurate illustration in the protologue (tab. 179a). The lectotype (MA; F neg. 29720: photos at GH, WIS) lacks flowers and fruit, but the isolectotype at P bears intact flowers. Specimen 8422/28 at G may be a type given that it is conspecific and a label on it indicates “Herb. Pavon,” but neither collection number nor locality are present. Other than the types, only a few collections of J. biflora have been seen among the hundreds of specimens studied at B, BH, BM, C, COL, COLO, CONN, G, F, GH, K, LD, MA, MO, NY, P, US, USM, VT, WIS. For this study of floral biology, seeds were collected at or near the type locality. Plants were grown at the University of Connecticut in a greenhouse with minimum evening temperatures of 7-10 C, and daytime temperatures of 15-29 C depending on cloud cover/sunshine; plants did not flower under warmer conditions present in other greenhouse rooms. Two fruits, collected in Peru, were orange at maturity and had 161 and 166 seeds each. Other observations were made in the greenhouse during April and May of 2000. The corolla (Fig. 1, 2), 1.3 cm long, is green and urceolate with a narrow mouth and broader base 1.4 cm in diameter. The limb (ultimately revolute) is made up of five narrowly triangular lobes alternating with inconspicuous lobules. The stamens and style are exserted beyond the mouth of the corolla on mature flowers. The anthers are purple before dehiscence. The calyx and corolla bear a dense indument of gland-tipped finger hairs. Stalked glands were present but rare on the abaxial face of the calyx. Neutral red stained both the multicellular head of these glands and the tip of the glandular finger hairs. Flowers remained open six to nine days (mean 7.25 days, n = 10 flowers), and remained open at night. The filaments and the style elongated during the first few days a flower was open, and only after a stamen elongated did its anther dehisce. During April, two to four of the anthers of a flower dehisced one day and the others dehisced the morning of the next day (n = six flowers). In May, however, when daytime temperatures in the greenhouse were noticeably warmer, all of the anthers of a flower dehisced in one day by 10:00 am (n = four flowers). The pistillate phase lasted two to three days (mean 2.3 days, n = 10 flowers) with the stigma protruding through the partially open corolla on the first day. This species is self-compatible. All three of the flowers manually emasculated and self-pollinated during the pistillate phase set fruit, as did all four of the flowers that were manually self-pollinated during the hermaphroditic phase. Many fruits were spontaneously set in the greenhouse in the absence of pollinators. These were orange at maturity, subspherical, and up to 14 mm in diameter. Nectar was sweet to the taste. A refractometer was used to estimate percent sugars from destructively sampled flowers. Average sugar concentration during the pistillate phase was 14.2 % (range 14 to 14.4, n = 2) and 38.9% for the hermaphroditic phase (range 26 to 57.6 percent, n = 5). Given that the pistillate phase precedes the hermaphroditic phase, and no nectar was removed by pollinators, the higher sugar concentration during the latter phase may have been due to continual nectar secretion and/or evaporation of water from the nectar. Flowers produced 0 to 47 µl of nectar (n = eight flowers). Nectar was clear at first, and although inconspicuous, could be seen through the corolla wall. Nectar became more conspicuous toward the end of the hermaphroditic phase: it turned amber to orange in color, as viewed both through the corolla wall and when removed with a clear pipette tip. |
| Greenhouse observations: Styles measured during the hermaphroditic phase ranged in length from 12 to 18 mm, with the extremes of this range present on the same plant at the same time. After two to three days of being pistillate, flowers with longer styles automatically self-pollinated because the stigma came in contact with the dehisced anthers. Flowers having shorter styles were similarly protogynous but were also herkogamous: dehisced anthers and the stigma remained separated by 3 mm and the stigma remained free of pollen in the absence of pollinators. Style length variation data are preliminary given low sample size (n = 7 measurements) and because for this study T.M. used three plants and did not know if these were from separate seeds, or cuttings of one or two individuals (seeds were sown in 1996 but these observations were not made until the year 2000). |
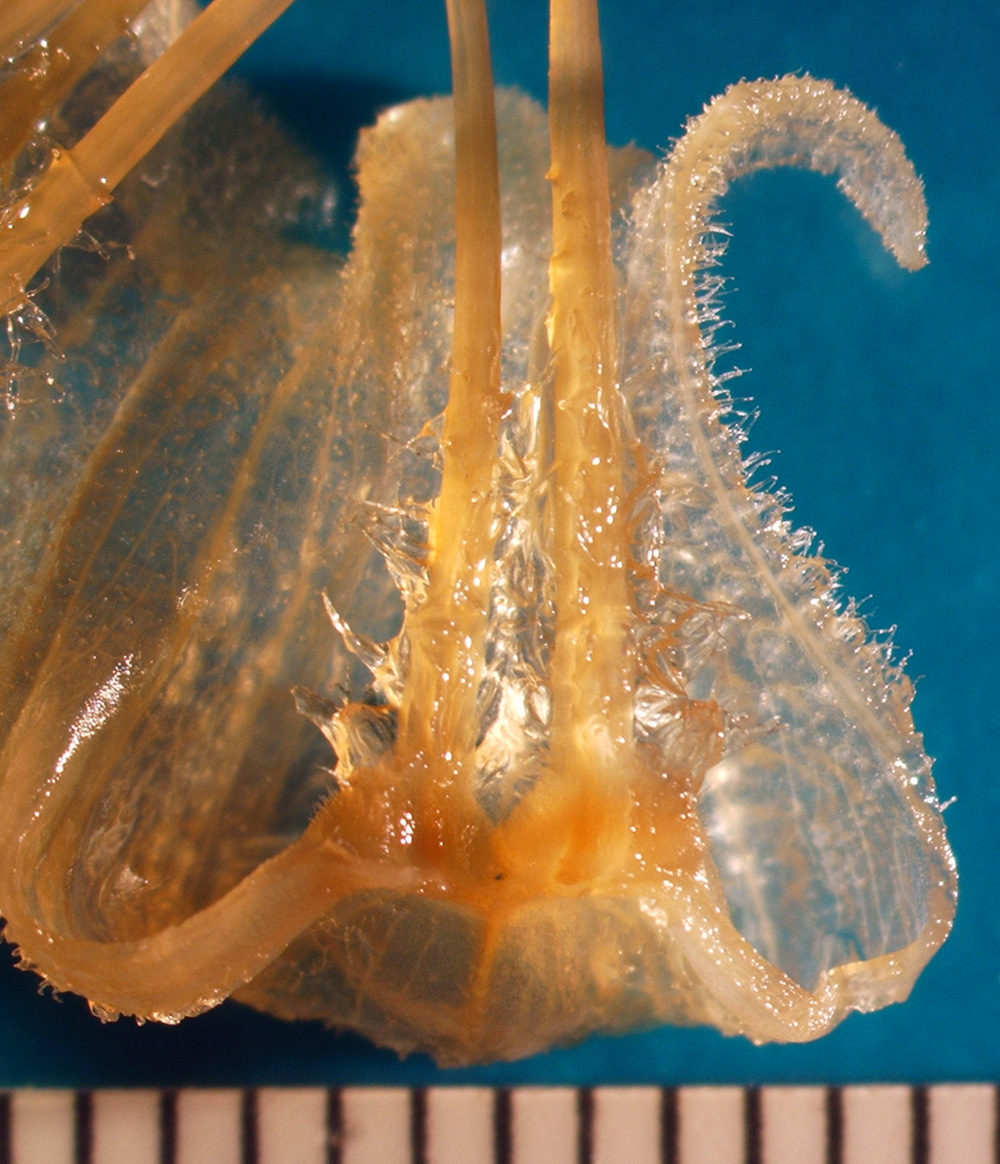 |
| Figure 5. Stamens (two) attached to base of corolla, showing laterally expanded base of stamen in ventral view, Jaltomata biflora; photo by Melissa Luna, Mione 608, flower from a plant grown in Connecticut and then preserved in 70% ethanol. |
 |
| Figure 6. Branches, leaves and inflorescenes of Jaltomata biflora. The corolla-androecium has dropped off of the flower on the right, revealing the calyx, yellowish ovarian disk and ovary. Photo by Daniel Mugaburu in Peru at or near type locality. |
 |
Figure 7. Stamens (two, Jaltomata biflora |
| In the The Journals of Hipólito Ruiz Ruiz wrote (page 113) "During the 11 months that we remained in the province of Tarma [one of the provinces of the department of Junin, Peru], exploring the area and carrying out botanical excursions in all directions, we discovered in those gorges and at the edge of the forest a considerable number of trees, shrubs, and herbs that had familiar uses, medicinal properties, or economic potential. The following are those that I collected, dried, and described; illustrations were done for nearly all of them." And then he lists numerous species, including (on page 114) Jaltomata biflora (as Saracha biflora). He makes no mention of human uses of this species in this work. |
