

photo by Thomas Mione
photo by Segundo Leiva G.
Jaltomata andersonii Mione |
Peru |
revised Oct 2022 |
The information on this page may be cited as a communication with professor Thomas Mione, Central Connecticut State University, Biology Department, Copernicus Hall, 1615 Stanley Street, New Britain, Connecticut 06050-4010, United States of America |
Rhodora 106: 118-23. 2004. |
Link to Jaltomata homepage |
The specific epithet was chosen to honor and thank Gregory J. Anderson, former doctoral advisor of the author. |
|
|
| Figure 1. Mione, Leiva G. & Yacher 620, photo by Thomas Mione |
Figure 2. Leiva G., Mione and Yacher 2127, photo by Segundo Leiva G. |
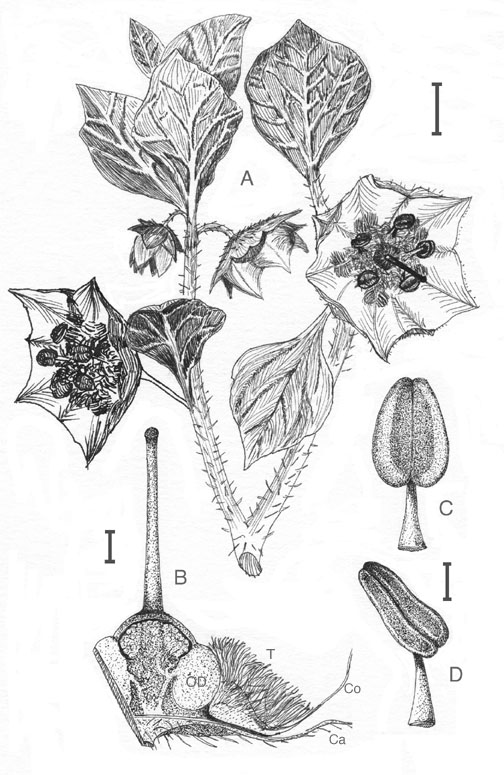 |
Figure 3. Jaltomata andersonii |
Jaltomata andersonii (Solanaceae) is distributed on the western slope of the Andes, in the Departments of Lima and Ancash, Peru, from 2300 to 3400 m of elevation. This species differs from others of the genus by having an unarticulated axis connecting the flower to the plant where all others have both a peduncle and a pedicel. The following combination of features also characterize this species: the hairs of leaves and axes are gland-tipped, the petiole is no longer than 1 cm, the flowers are solitary, the corolla is broadly crateriform-rotate, purple and up to 3.7 cm in diameter, and the filaments are extremely villous at their base. This species was discovered during fieldwork by Segundo Leiva G., Thomas Mione and Leon Yacher in 1998. Mione and Coe (1992) placed the following binomials in synonymy with Jaltomata aspera (Ruiz & Pav.) Mione: Saracha ciliata Miers, Saracha lacrima-virginis Bitter, and Saracha urbaniana Bitter & Dammer. The type specimens of all of these binomials have a single relatively large flower per inflorescence, and were collected either on the west slope of the Andes or in the lomas [a fog-fed desert habitat of the west coast of South America having a high level of endemism (Dillon 1997)]. The species represented on this web page also has a single flower per inflorescence and is from the west slope of the Andes. Given these similarities, one may wonder if one of the aforementioned binomials represents the earliest name of J. andersonii. None of the above binomials represent the earliest name of this species for the following reasons. The type specimens of the above Saracha binomials have both a peduncle and a pedicel, and were described as producing red nectar. According to the protologue of Jaltomata aspera (Ruiz & Pavón 1799) the type was collected in Amancaes and the flower is violet in the middle ("in centro violacea"). Amancaes is located within lomas habitat, and is within several hundred meters of sea-level [according to a specimen label of J. aspera collected at Amancaes (Weberbauer 5246a, US), and Dillon, Field Museum of Chicago, personal communication]. The description of the flower as violet in the middle suggests that the type produced red floral nectar, and red floral nectar has been documented for J. aspera (Vilcapoma S. and Granda P., Universidad Nacional Agraria La Molina, personal communication). In contrast, J. andersonii has a single unarticulated axis joining the flower to the plant (Figure 3A), lacks red nectar, and grows at high elevations, not in the lomas habitat. Thus, all of the above Saracha binomials are synonyms of J. aspera, as originally reported (Mione and Coe 1992). |
| Character | Description | Figures on this page |
Habit & Height |
Much branching, presumably perennial, herb to 20 cm | |
Branches, young |
pilose, the hairs long, soft, finger-type, gland-tipped | |
older |
||
Leaves, size |
Leaves alternate, sometimes geminate, the blades 4-7.5 cm long (to 10.5 cm on Spooner et al. 7364a ) X 2-3.5 cm wide, 3-4 pairs of primary veins, the margin nearly entire to sinuate-dentate | |
| shape | the apex often somewhat acuminate, the base usually cuneate and symmetrical or nearly so, the blades ovate | |
| hairs | adaxially densely pilose due to erect finger hairs, abaxially sparsely pubescent (nearly glabrous on Laudeman 5365), the margin ciliate, | |
| petiole | to 1 cm long | |
Inflorescence |
flowers solitary | |
peduncle |
unarticulated, villous, 13-18 mm long | |
pedicel |
||
Calyx when flowering |
Calyx green, rotate, 16-21 mm across at anthesis, the lobes triangular with an acute apex, abaxially with a raised costa, bearing gland-tipped finger hairs to 3 mm long, and stalked multicellular glands (illustrated in Mione and Serazo 1999) 85-100 µm long, adaxially with a uniform covering of erect, glandless hairs 0.24 - 0.3 mm long, the margin with two classes of hairs: gland-tipped finger hairs to 2.8 mm long and glandless hairs to 0.1 mm long; calyx to 25 mm across with fruit. | |
| shape / position when flowering | ||
| at fruit maturity | ||
| hairs | ||
Corolla color |
purple with 5 pairs of dark-green maculae in a proximal ring, each pair of maculae straddling the main vein that extends radially to the tip of a corolla lobe, closing for the night | 1, 2 |
shape and size |
broadly crateriform-rotate, to 3.7 cm in diameter, | 1, 2, 3A |
lobes/lobules |
5-lobed | |
hairs
|
the margin ciliate, with glandless finger hairs to 0.6 mm long; both faces of corolla with abundant, evenly distributed stalked multicellular glands 85 - 100 µm long. | |
Stamen length including anther |
5, 6.9-7.2 mm long, filaments straight, extremely villous at base |
3A, B |
| length stamens exserted beyond distal end of corolla (applicable if corolla has a well-defined tube) | ||
| no | Mione et al. 620 was observed with a dissecting microscope Sept 2017 | |
| base expanded laterally? | ||
| filaments | ||
| anther color | drying brown, or yellow if covered with pollen. | 3C & D |
| anther size | Anthers 1.9-2.8 mm on pressed specimens, to 3.9 mm long on specimens preserved in alcohol in Peru | 3C & D, 4 |
| anther mucronate/mucronulate | no | 4 |
| insertion of filament into anther | filament is inserted on the lower ventral face | 4 |
| anthers of a flower open simultaneously? | no data |
|
| pollen quantity per flower | 467,000 (n = 1 flower, Mione et al. 616) | count by Emmett P. Varricchio 2013 |
| pollen grain size | ||
| corona | no | |
Stigma |
Truncate: the stigma the same width as the distal end of the style, 0.21-0.3 mm wide (pressed), 0.36 mm wide (preserved in alcohol in Peru), both measured perpendicular to the length. | 3B |
Style |
Style straight, 5.7-6.3 mm long, 0.15-0.36 mm wide at midlength. Gynoecium glabrous, except for stigma papillae to 13 µm long; | 3B |
Ovary |
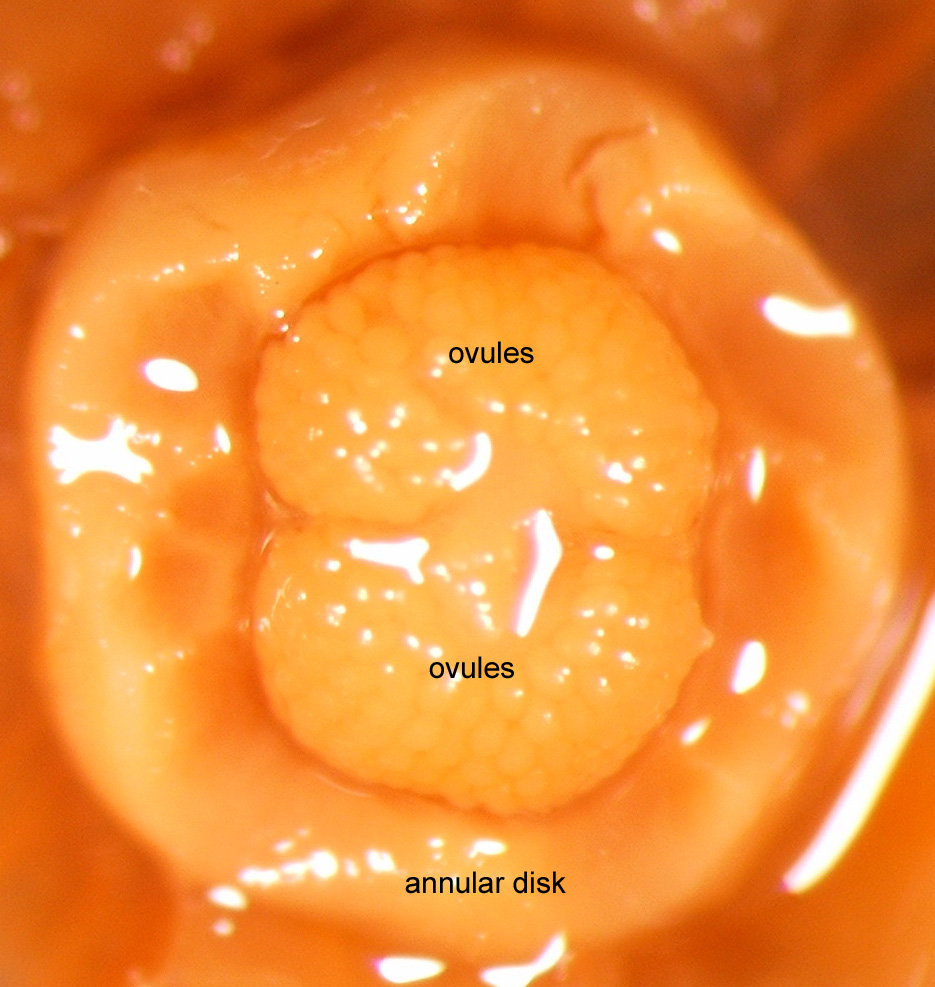
Ovary glabrous but appearing villous because of the dense vestiture of the stamen bases. Ovarian disk broad, girdling base of the ovary | 3B, 5 |
Ovules per ovary |
226 (n = 1 flower, Mione et al. 616) | Count by Thomas Mione Fig. 5 |
Nectar |
||
| Herkogamy | ||
| Protogyny | ||
Fruit color (at maturity) and size |
Berries of unknown color, probably orange or red, 12 mm across (measured on Laudeman 5365 ). | |
Seeds per fruit |
no data |
no data |
Seed Size |
no data |
no data |
Chromosome number |
no data |
no data |
Growability in Connecticut, USA |
no data |
no data |
How long does it take from flower to ripe fruit? |
no data |
no data |
Self-Compatible? |
no data |
no data |
Seed Germination |
no data |
no data |
Ratio of pollen to ovules |
2,066 (467,000 / 226 ovules; Mione et al. 616) | 2013, n = 1 flower collected in Peru (not grown for study) |
| Character | Description of Jaltomata andersonii | Figures on this web page |
Geographic and Altitudinal Distribution. Peru, mostly department Lima, one specimen from department Ancash.
| Department | Province | Locality | elevation m |
habitat | date |
collector (herbarium) | Data Entry |
| Ancash | Bolognesi | Chiquián | 3,150-3,400 |
cultivated grounds below the village, with Agave americana and Mutisia dominant | 5 Feb-2 Apr 1997 | M. Weigend & N. Dostert 97/173 (F) | Jan 2007 |
| Lima | Cajatambo | Baños de Churin | 2,591 |
among rocks | Feb 1946 | C. Laudeman 5365 (K) | Jan 2007 |
| Lima | Canta | Cuesta de Huamantanga | 2,720 |
10.8 km NW from the Lima to Canta road on the road to Huamantanga | 7 Apr 1999 | D. Spooner, A. Salas, R. Torres & R. Hoekstra 7364a (WIS) | Jan 2007 |
| Lima | Huarochirí | Lima to La Oroya highway, at km 73 walk up | 2,300 |
dry treeless hillside, sandy soil with rocks, with Cactaceae, Carica candicans, Lycianthes sp., and Schinus molle | 15 Jan 1998 | T. Mione, S. Leiva G. & L. Yacher 616 (NY), S. Leiva G., T. Mione & L. Yacher 2122 (F, HAO) | Jan 2007 |
| Lima | Huarochirí | Lima to La Oroya highway, km 83 | 2,550 |
roadside with Agave | 16 Jan 1998 | T. Mione, S. Leiva G. & L. Yacher 620 (CONN), S. Leiva G., T. Mione & L. Yacher 2127 (HAO) | Jan 2007 |
| Lima | Huarochirí | highway from Lima to La Oroya between km 82 & 83 |
2,500 |
with J. propinqua (Miers) Mione & M. Nee | 16 Jan 1998 | TYPE SPECIMEN T. Mione, S. Leiva G. & L. Yacher 622 (Holotype: NY!; Isotype: CONN!). |
Jan 2007 |
Jaltomata andersonii grows in treeless, usually dry, rocky habitats, and flowers in January, February and March. Jaltomata andersonii is most similar to J. quipuscoae. Both species have one relatively large flower per inflorescence, a purple corolla, and are much-branching, low-growing perennial herbs that grow in Peru. The similarity of these species is superficial: these two species are classified in two different sections of the genus (J. andersonii in section Hebecladus, and J. quipuscoae in section Modillonia). Jaltomata quipuscoae has both a pedicel and a peduncle, filaments are nearly glabrous, red floral nectar is produced, and it grows from 300 to 860 m and 2607 to 2625 m of elevation. In contrast, J. andersonii has an unarticulated axis (by definition a peduncle, though the peduncle may have been lost) connecting the flower to the plant, the filaments are extremely villous at their base, red nectar is not produced, and grows at elevations from 2300 to 3400 m. The hairs of J. andersonii are soft to the touch and gland-tipped (as observed on living plants of Mione et al. 616, 620 and 622). The only specimen of J. andersonii collected in Department Ancash (Weigend & Dosert 97/173) had "brown and whitish green" (not purple) flowers and grows at a higher altitude than all other specimens. Study of this specimen again (January of 2007) revealed a single unarticulated axis instead of both a peduncle and pedicel, and extremely hairy filaments at base, just like the other specimens of J. andersonii. Leaf shape and size for this specimen matches the others. Thomas Mione, Segundo Leiva G. and Leon Yacher were able to see both J. aspera and J. andersonii in bloom during January of 1998 because of the rains associated with the El Niño event of 1997/1998. We did not interview local people about uses, but the fruits of J. andersonii may be eaten, given that ripe fruits of most other South American species of this genus are consumed. |
 |
Figure 4. Two anthers, both undehisced, from the Ventral face of
Mione, Leiva G. & Yacher 616 Photo by Thomas Mione Anthers had been Units, vertically
|
Figure 5. Ovary Of |
 |