Jaltomata aijana Mione & S. Leiva
|
Peru |
revised January 2022 |
|
The information on this page may be cited as a communication with professor Thomas Mione, Central Connecticut State University, Biology Department, Copernicus Hall, 1615 Stanley Street, New Britain, Connecticut 06050-4010, United States of America. |
|
|
Link to the Jaltomata of Ancash, Peru
|
|
Replaced synonym: Hebecladus weberbaueri Dammer, Bot. Jahrb. Syst. 37(5): 638. 1906
[not J. weberbaueri (Dammer) Mione, Novon 2(4): 383. 1992; Basionym: Saracha weberbaueri Dammer, Bot. Jahrb. Syst. 37(5): 638. 1906].
This taxonomic revision corrects the erroneous placement of H. weberbaueri Dammer in synonymy with J. bicolor (Mione et al. 1993). |
 |
Figure 1. Jaltomata aijana. A—Flower in pistillate phase, anthers are purple. B—Flower near center has two dehisced and three undehisced anthers showing nonsimultaneous anther dehiscence. The flower on the left opened more recently; we can infer this because the stamens are not yet exserted.
C—Corolla was opened longitudinally to show androecium and gynoecium; two of the five stamens were removed. D—Distal end of flower, showing alternating lobes and lobules (10 total) and dehisced anthers. E—Woody stem, smallest units are mm. F—Calyx in overhead view, attached to fruit hidden under the calyx, units along bottom are mm.
All scale bars equal one cm. Photos are of Mione et al. 863 = Leiva G. et al. 5892 except A is Mione et al. 728.
Photos were taken by T. M. in the field except B was taken by S. L. G. in the field. |
 |
Figure 2. Ripe fruits of Jaltomata aijana. Plant grown in CCSU greenhouse. Units are mm. One of the fruits measured 8.7 high by 13.5 wide, and the other 8.9 wide by x 14 mm wide. Mione et al. 872 |
|
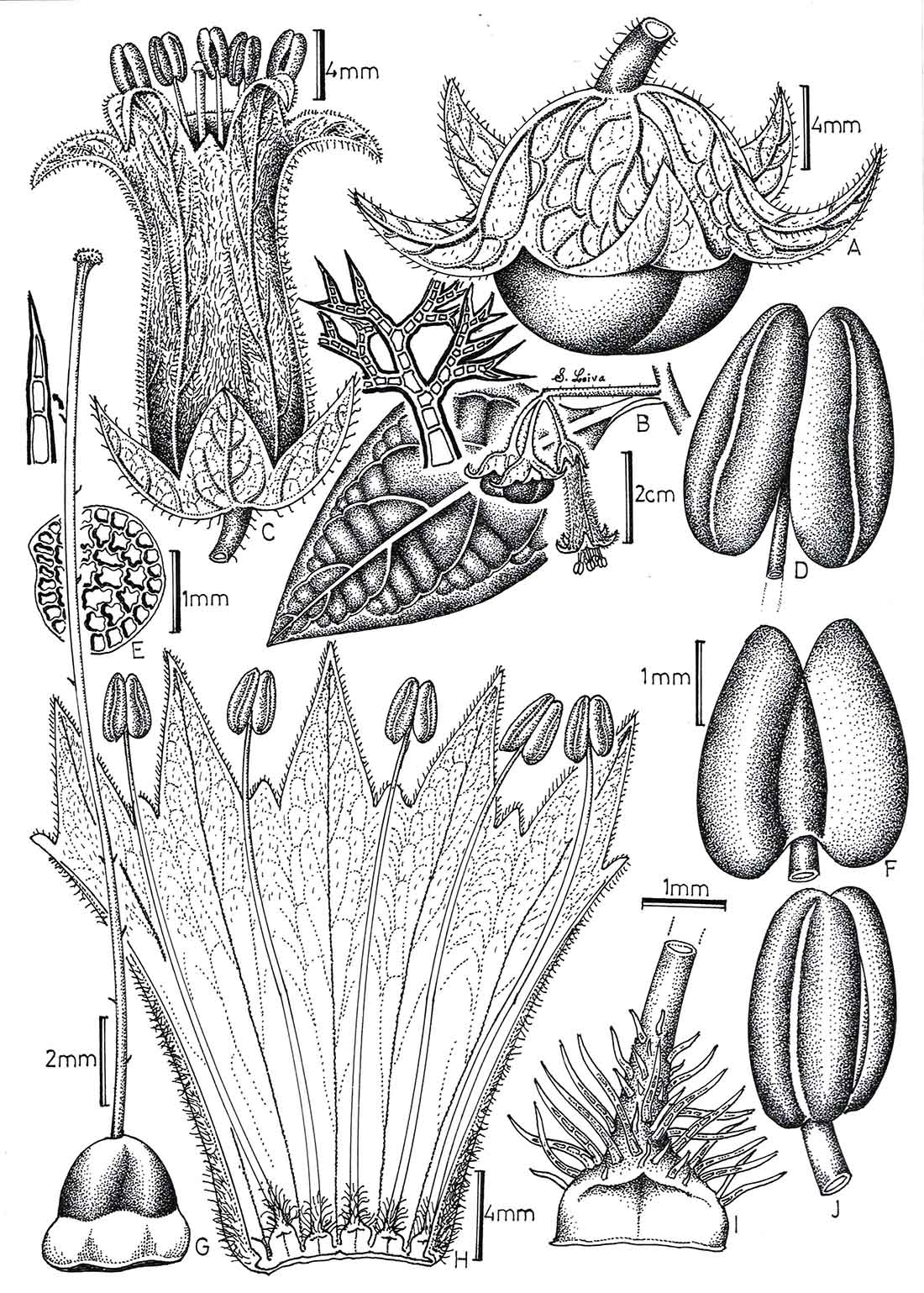
Figure 3. Jaltomata aijana. A—Fruit in side-view, approximately half-hidden by calyx.
B—Branch with inflorescence and leaf with an expanded view of a dendritic hair.
C—Open flower.
D—Anther in ventral view.
E—Seed.
F—Anther in dorsal view.
G—Gynoecium.
H—Corolla with adnate stamens dissected open longitudinally.
I—Base of staminal filament in ventral view.
J—Anther in lateral view.
Illustration by Segundo Leiva González, S. Leiva G. et al. 5892 |
 |
| above, Figure 4. Flowers, leaves, Jaltomata aijana, numbered units are cm, photo by Thomas Mione in Peru, Mione et al. 863 |
Description of Jaltomata aijana based on numerous herbarium specimens from throughout the range of the species:
| Character |
Description |
Figures on this page |
Habit & Height |
Shrub to 2 (–3 m) high. |
|
Branches, young |
younger stems 4- or 5-sided, purple on upper surfaces, green on lower surfaces, sparsely to densely pubescent with eglandular unpigmented mostly simple hairs |
|
| older |
brown, terete, with lenticels, glabrous, to 15 mm diameter at the base |
|
Leaves, size |
Basal leaves alternate, the distal geminate; the blade ovate, to 8 (–14.5) cm long X 5 (–12) cm wide, the younger membranous, the older somewhat coriaceous, the upper face sparsely pubescent to glabrous, the lower puberulent to glabrous, the apex usually acute, |
|
| shape |
the base of smaller leaves at times cuneate, the base of the largest leaves truncate, the margin usually entire but the largest are sinuate to sinuate-dentate; Leaves single, paired or verticillate; usually ovate but variable in size, shape and margin. |
|
| hairs |
glabrate to sparsely pubescent with finger-like and dendritic hairs;
cultivated plants of Mione et al. 872 had leaves that were villous, the hairs all gland-tipped, to 2 mm long, with notable and obvious length variation among hairs of the leaves, some finger hairs twice as long as others; a few hairs were forked and gland-tipped at both termini |
hypocotyl and cotyledons of seedlings are pubescent with unpigmented conspicuously gland-tipped finger hairs
(Oct 2016, Mione et al. 872) |
| petiole |
the petiole to 6 mm (–3 cm) long, with a conspicuous main vein abaxially |
|
Inflorescence |
flowers 2–5 (–6) per inflorescence |
up to 10 flowers per inflorescence (if one includes small flower buds) on cultivated plants of Mione et al. 872 |
| peduncle |
green, terete, sparsely pubescent, to 3.1 cm long |
|
| pedicel |
purple, terete, sparsely pubescent, to 1.4 cm long |
|
Calyx during anthesis |
abaxially green to purple, five-lobed (stellate in outline), 1.6 cm diameter, sparsely pubescent with simple to dendritic eglandular hairs, the lobes triangular, to 6.5 mm long X 5 mm wide, the main vein somewhat raised abaxially. |
|
| shape / position when flowering |
planar to recurved (Figure 1A, B) |
|
| at fruit maturity |
enclosing 1/3 to 1/2 of the fruit (Figure 2A),
to 3.1 cm diameter; 15 mm from pedicel to lobe tip |
|
Corolla color |
Corolla green |
|
| green spots |
no |
|
| purple ring |
no |
|
| purple in base of corolla |
yes: at least some collections having purple at the base |
Figure 11 |
| shape and size |
tubular, the tube and limb together to 3.5 cm long, the tube narrowing slightly from proximal to distal, 6–10 mm diameter at midlength |
|
| lobes / lobules |
10 total: the lobes alternate with the inconspicuous lobules. |
|
| hairs |
the exterior of the corolla densely pilose, the hairs simple or sometimes dendritic, the interior of the tube glabrous, the limb pubescent on both faces |
|
Stamen length including anther |
5, adnate to base of corolla, connivent, hidden inside corolla tube when corolla opens, elongating at unequal rates but finally equal, 3.1–3.5 cm long including anther, exserted beyond mouth of corolla |
|
| length stamens exserted beyond distal end of corolla (applicable if corolla has a well-defined tube) |
0 - 8 mm in Department Ancash |
|
| radial thickenings |
no
|
726, 728, 872 were dissected carefully |
| base expanded laterally? |
yes |
Figure 2I and dissection microscope observations by Mione September 2017 |
| filaments |
pale-green, the proximal 10% villous with unpigmented, eglandular, simple hairs
|
|
| anther color |
anthers dark purple, the connective greenish |
Figures 1A, 1C, 13 |
| anther size |
sagittate after dehiscence, glabrous, conspicuously longer than wide, 3-3.5 mm long X 1.8-2 mm wide (fresh), 2.5-3 mm long X 1-1.5 mm wide (pressed specimens), finally in close proximity with stigma or to 2 mm beyond stigma |
|
| anther mucronate? |
yes: mucronulate
(not
showing in Figure 2) |
dissecting microcope observations by Mione on fresh anthers from cultivated plant of Mione et al. 872 |
| insertion of filament into anther |
directly under anther |
Figure 13 |
| anthers of a flower with temporally staggered dehiscence? |
yes |
Figure 1B |
| pollen grain size |
|
|
| corona? |
no |
|
Stigma |
capitate (Figure 1C, 2G), dark green, exserted, shallowly bilobed, 0.5–1 mm diameter (pressed specimens) |
|
| Style |
straight, filiform, pale-green, lighter than the stigma, 2.3–2.9 cm long |
|
| Ovary |
2.4 mm high X 3.4 mm wide including the annular disk, containing 236 ovules, the annular disk pentagonal, pale-orange and approximately 60 % of the height of the ovary |
|
Nectar |
Up to 30 microliters per flower, flowers from which nectar had not been previously removed (cultivated plants in greenhouse, Mione et al. 872)
Probably translucent most of the time in the field (in the wild) because during field collections in Peru I never noticed colored nectar. However, pale-orange during both pistillate and hermaphroditic phases (Jamie Kostyun, personal communication May 2013, cultivated plants) |
|
| Herkogamy |
|
|
| Protogyny |
yes |
Mione 872, cultivated |
| Fruit color (at maturity) |
Orange
|
Figure 2 |
| Fruit Size and Seeds per fruit |
9 mm high X 13.5 mm wide containing 82 seeds (Mione et al. 872);
11 mm (pole to pole) X 13 mm diam not ripe but may represent full size (Mione et al.863);
22, 30, 51 seeds per fruit (Mione et al. 863)
206 seeds per fruit
Data from the field is not comparable with data from the greenhouse: for example,
J. aijana collection Mione et al. 872 had 82 seeds in one fruit in the field (year 2016), and 38 to 40 seeds per full-size, ripe fruit (weeks after manual self-pollinations) in the greenhouse (year 2018).
|
Berry globose, compressed at the poles, to 11 mm (pole to pole) X 13 mm diameter, the style persistent |
| Seeds |
brown, sub-orbicular to sub-triangular, the surface reticulate (Fig. 2E),
1.55–1.7 X 1.25–1.4 X 0.5–0.6 mm thick |
|
| Mione's collection numbers of this species |
625, 627, 726, 728, 863, 870, 871, 872 |
|
For other descriptions see |
protologue and description of Hebecladus weberbaueri Dammer in Macbride (1962) |
|
| Character, Jaltomata aijana |
|
|
Growability in Connecticut, USA |
Will grow and flower in a
greenhouse, and in a window of an air conditioned building, in Connecticut |
Mione et al. 872 |
How long does it take from flower to ripe fruit? |
5 weeks and 2 days (two fruits, greenhouse) |
Mione et al. 872 cultivated at CCSU |
How long does it take from seed germination
until flowering? |
about 6 months: 20 October 2016 to 15 April 2017 |
Mione et al. 872 cultivated at CCSU |
Flowers Closing For The Night? |
No |
Mione 872 cultivated at CCSU |
Self-Compatible? |
Yes
|
Mione et al. 872 at CCSU |
Seed Germination |
26 days from planting to first above-ground signs of germination, seeds in cup of potting mix on warm mat, no GA |
Mione et al. 872 |
| Pollen quantity per flower |
65,450 and 79,063 Mione et al. 728, n = 2 flowers
93,250 Mione et al. 726, n = 1 flower |
flowers collected in Peru (not grown for study), counts by Elisabeth dos Santos &
Emmett P. Varricchio |
Ovules per ovary |
236 Mione et al. 728, n = 1 flower |
count by Thomas Mione, the value shown at left is double the count from one locule |
Ratio of pollen to ovules |
277 (65,450 / 236 ovules) |
Mione et al. 728, n = 1 flower |
Chromosome number |
no data |
|
 |
 |
| above, Figure 5. A flower bud was pulled open longitudinally to allow view of stamens. Anthers are dark purple. Given that stamens of open flowers of this collection were 31 to 35.5 mm long, and filaments in this photo are approx 1 mm long, we can conclude that filaments elongate tremendously between the floral bud stage (showing) and the flower being functional. Photo by Thomas Mione in the field in Peru, Mione et al. 863. |
above, Figure 6. Flower with petal removed to show stamens and style, Jaltomata aijana, photo by Thomas Mione in Peru, Mione et al. 863 |
 |
 |
| abaove, Figure 7. Distal end of flower, Jaltomata aijana, photo by Thomas Mione in Peru, Mione et al. 863 |
above, Figure 8. Flower opened recently, probably within the last few hours (we can infer this because the stamens have not yet elongated enough for us to see them in a view perpendicular to the corolla tube). Photo by T. Mione in Peru, Mione et al. 863. |
 |
 |
| Figure 9. Note the orange nectar inside the corolla tube, evident only in cultivated plants, not seen in the wild. Plants grown by and photo by Jamie Kostyun, Mione et al. 726. |
Figure 10. Note the orange nectar inside the corolla tube, evident only in cultivated plants, not seen in the wild. Plants grown by and photo by Jamie Kostyun, Mione et al. 726. |
We collected specimens of the genus Jaltomata in Peru in 1998, 1999, 2005, 2007, 2008, 2010, 2011, 2013, 2015 and 2016. Specimens have been deposited at CONN and HAO. Herbarium specimens were borrowed from or studied at BH, CONN, F, G, GH, K, M, MA, MO, MOL, NY, P, US, USM, VT and WIS with herbarium acronyms as in Thiers (continuously updated). Seeds of Mione et al. 872 were measured and Leiva González et al. 5892 was used for the seed count. Other characters were measured on all of the specimens we collected in Department Ancash (Table 1).
Phenology
Jaltomata aijana flowers November through June. Flowers are protogynous: both open flowers having undehisced anthers (pistillate phase) and open flowers having dehisced anthers (hermaphroditic phase) were seen on the same plant at the same time (Figure 1B) both in the field on plants cultivated in Connecticut, USA. Anther dehiscence involves anther color change from purple (undehisced) to greenish (dehisced), and is temporally staggered within a flower (Figure 1B). This species is self-compatible and flowers do not close for the night (observations made on plants cultivated in Connecticut, USA). In the greenhouse, during February of 2020 I tagged a flower to observe it daily: The stigma protrudes from the closed corolla for one day; the following day the corolla is open but the anthers remained undehisced (pistillate phase); the following day all anthers dehisced by 9 am (temperature 17.5 C) and the flower remained functionally hermaphroditic for the remainder of this day and the next; the following day the corolla-androecium abscised by 10 am. From pollination (by hand to record the date) to ripe fruit takes 5 weeks and 2 days (Mione et al. 872, 2 fruits 20 Jan to 26 Feb 2018, and one fruit 27 Jan to 4 March 2020).
Habitat
The habitat of Jaltomata aijana is open and shrub-covered hillsides, grazed rocky slopes, moist ravines, crevices in stone walls, along edge of agricultural field where it meets a river, roadsides. It lives in association with plants of Eucalyptus amygdalina Labill. “eucalipto” (Myrtaceae), Bidens pilosa L. “cadillo,” Viguiera weberbaueri S. F. Blake “suncho,” Ambrosia peruviana Willd. “marco,” Tagetes minuta L. “chiche,” Mutisia acuminata R. & P. (Asteraceae), Jaltomata weberbaueri (Dammer) Mione “frutilla,” Brugmansia sanguinea (R. & P.) D. Don “floripondio rojo” (Solanaceae), Pisum sativum L. “arveja,” Otholobium pubescens (Poir) J. W. Grimes “culén” (Fabaceae), Rumex crispus L. “acelga” (Polygonaceae), Alnus acuminata Kunth “aliso” (Betulaceae), Passiflora tripartita (Juss.) Poir. “puro puro” (Passifloraceae), Sambucus peruviana Kunth “saúco” (Adoxaceae), Alonsoa meridionalis (L. f.) Kuntze (Scrophulariaceae), and the genera Urtica L. (Urticaceae), Cestrum L., Solanum L., Salpichroa Miers (Solanaceae), Calceolaria L. (Calceolariaceae), Barnadesia Mutis ex L. f. (Asteraceae) among others.
Phylogeny
Jaltomata aijana was included in the molecular phylogeny of Miller et al. (2011) as “J. bicolor 726” where it was found to be most closely related to J. umbellata (R. & P.) Mione & M. Nee in both the strict consensus and the majority-rule consensus trees. We now understand that the circumscription of J. bicolor was artificial when Miller et al. (2011) published, and that J. bicolor and J. aijana are distinct and defendable species (Table 2).
Local names
Table 1. All specimens of Jaltomata aijana examined, all collected in Department Ancash, Peru.
Province. Locality |
elevation
m |
habitat |
date |
collector (herbarium) |
See footnote 1. |
3200– 3400 |
interwoven grasses and bushes ("graminosis fruticibus intertextis")2 |
27 March 1903 |
Type of Hebecladus weberbaueri Dammer.
Weberbauer 2652: Isotype G!, photos of B specimen (destroyed) GH!, NY! |
Huari. road from San Marcos to Chavín de Huántar. Río Mosna valley near Olayan |
2800 |
Low but closed tropical dry forest vegetation with Arnaldoa, Llangunoa |
13 Mar 2001 |
M. Weigend et al. 5151 (M) |
Huaraz. km 186 Recuay - Huaraz |
3150 |
on dry ground along bank of tributary to Río Santa |
Jan 1998 |
T. Mione, S. Leiva G. & L. Yacher 627 |
Huaraz. 15 km S of Huaraz, 77 29' W, 9 41' S |
3450 |
brush borders in agricultural land |
27 Jan 1985 |
D. N. Smith et al. 9388 (NY) |
Bolognesi. km 107, poblado Sta. Rosa (ruta Pativilca-Recuay) |
3300 |
borde de río entre Ambrosia sp., Barnadesia |
18 Jan 1998 |
S. Leiva, T. Mione & L. Yacher 2133 (F)
T. Mione, S. Leiva G. & L. Yacher 625 |
Bolognesi. Huacacorral, 22.4 km E of Conococha, 77.2 W, 10.1 S |
3820 |
growing among Opuntia in clay soil |
3 Apr 1999 |
D. M. Spooner et al. 7356a (WIS) |
Bolognesi. 9 57' 23.5" S, 77 05' 57.0" W |
3971 |
steep roadside |
20 May 2016 |
T. Mione, S. Leiva G. & L. Yacher 870
S. Leiva, T. Mione & L. Yacher 6048 |
Bolognesi. Arriba de Chiquián |
3500–3600 |
falda pedregosa |
31 Mar 1957 |
R. Ferreyra & E. Cerrate 12133 (USM) |
Bolognesi. Chiquián |
3400–3900 |
fields, pastures and quebrada above village |
5 Feb–2 Apr 1997 |
M. Weigend & N. Dostert 97/185 (F) |
Bolognesi. Below Chiquián
10 08' 16.3" S, 77 09' 43.2" W |
3338 |
roadside |
20 May 2016 |
T. Mione, S. Leiva G. & L. Yacher 871
S. Leiva, T. Mione & L. Yacher 6049 |
Bolognesi. Chiquián |
3338 |
hedgerows, trail edges |
20 May 2016 |
T. Mione, S. Leiva G. & L. Yacher 872
not pressed, seeds only |
Bolognesi. highway Pativilca - Recuay,
10 9' 47" S, 77 20' 16" W |
3645 |
steep roadside |
15 Jun 2005 |
T. Mione, S. Leiva G. & L. Yacher 728
digital photos only, not pressed |
Recuay. Bosque de Pararín |
2800– 3000 |
bosque |
24 May 1988 |
N. Valencia 2168 (NY) |
Recuay. km 110 road from Lima to Huaraz, 19.7 km N of Cajacay, 77.3 W, 10.2 S |
3600 |
growing among Opuntia and shrubs |
3 Apr 1999 |
D. M. Spooner et al. 7359a (BH, P, WIS) |
Recuay. km 107 between Recuay and Pativilca |
3300 |
moist ravines |
30 Jan 1983 |
M. Dillon et al. 3178 (BH, F) |
Recuay. ca. Laguna Conococha, km 107 (Pativilca-Recuay) |
3500 |
quebrada de arbustos |
17 Nov 1995 |
S. Leiva G. 1738 (HAO) |
Recuay. Culquimarca (Pativilca-Conococha) |
3600 |
en ladera de arbustos |
27 May 1970 |
A. López M. et al. 7601 (US) |
Aija. En la bajada de Tranca hacia Huayán |
3000–3200 |
no data |
23 Apr 1983 |
C. Ochoa & A. Salas 15,168 (F) |
Aija. 7 km E of town of Aija, canyon San Juan (containing Río San Juan), at Cuesta de Melliso, 77.6 W, 9.8 S |
2945 |
on a rocky slope with grasses, Calceolaria, Solanum hirsutum |
1 April 1999 |
D. M. Spooner et al. 7352c (WIS) |
Aija. 6 km down from Aija
9 48' 18" S, 77 36' 18" W |
3309 |
along river and edge of agricultural field |
14 Jun 2005 |
T. Mione, S. Leiva G. & L. Yacher 726 |
Aija. Road from Aija to Huarmey |
3474 |
roadside |
20 May 2015 |
T. Mione, S. Leiva G. & L. Yacher 863
S. Leiva G., T. Mione & L. Yacher 5892 (HAO) |
1. In the protologue the type locality is given as “Prov. Cajatambo, dep. Ancachs: Ocros 3200-3400 m.” On modern maps Ocros is the capital town of the province Ocros, at the south end of the department of Ancash, while province Cajatambo is at the north end of the department of Lima, just south of the department of Ancash.
2. from the protologue.
 |
above, Figure 11. Two corollas of Jaltomata aijana, removed from flowers and inverted to show purple pigmentation in base of corolla.
Photo by Thomas Mione in Peru, Mione et al. 863. |
Table 2. Comparison of Jaltomata species most similar to J. aijana. All three species are shrubs having 10-lobed corollas (the lobes and lobules alternating),
purple anthers, unpigmented floral nectar (in the field) and orange fruits.
character |
J. aijana Mione & S. Leiva |
J. bicolor (R. & P.) Mione |
J. biflora (R. & P.) Benítez |
Distribution
|
Peru: Department Ancash |
Peru: Departments Lima, Huancavalica, and Moquegua |
Peru: Department Junín |
Altitude (m) |
2800–3970 |
2100–3800 |
2700–3200 |
Corolla color |
green |
proximal 2/3 intensely purple, distal 1/3 green |
green |
Corolla shape |
straight-tubular |
urceolate-tubular |
urceolate |
Indument of external face of corolla tube |
unpigmented |
purple |
unpigmented |
Radial thickenings in corolla1 |
no |
no |
yes |
Nectar (floral) color when plants are cultivated |
clear, turning orange with age2 |
clear |
clear, turning orange with age3 |
Fruit, at maturity, hidden by calyx? |
upper ¼ to 1/2 of fruit hidden by calyx |
no |
upper ¼ to 1/3 of fruit hidden by calyx |
Fruit size (mm)(measured in the field, not cultivated) |
9 x 13.5
11 x 13 |
8.5 x 18 |
11 x 16 |
Seeds per fruit (fruits collected in the field) |
206 |
196, 216 |
119, 161, 166 |
Fruits eaten by humans |
no data |
Yes4 |
Yes4 |
Synonyms and page number(s) in Macbride (1962) |
Hebecladus weberbaueri Dammer, p. 40 |
Hebecladus bicolor (R. & P.) Miers, p. 30; Atropa biflora R. & P., p. 30; H. intermedius Miers, p. 35. |
Saracha biflora R. & P., p. 31 |
Author's collection numbers |
625, 627, 726, 728, 863, 870, 871, 872 |
612, 617, 795, 880 |
608, 876 |
1. A subset of species of section Hebecladus have radially oriented thickenings that extend from the base of each stamen toward the corolla lobule (or where
the lobule would be if present); where present, these create troughs (between radial thickenings) that hold nectar at the base of the corolla.
2. Nectar turning orange with age has been observed in the CCSU greenhouse on plants grown by T.M., and by Jamie Kostyun, personal communication (2013), from observations on cultivated plants.
3. Mione et al., (2001).
4. Leiva González et al., (2016a, pp. 49–50) and fieldwork by the authors.
 |
| Figure 12. Jaltomata aijana in Department Ancash, Peru. Leiva G., Mione & Yacher 5892 = Mione, Leiva G. & Yacher 863, Photo by Segundo Leiva G. |
 |
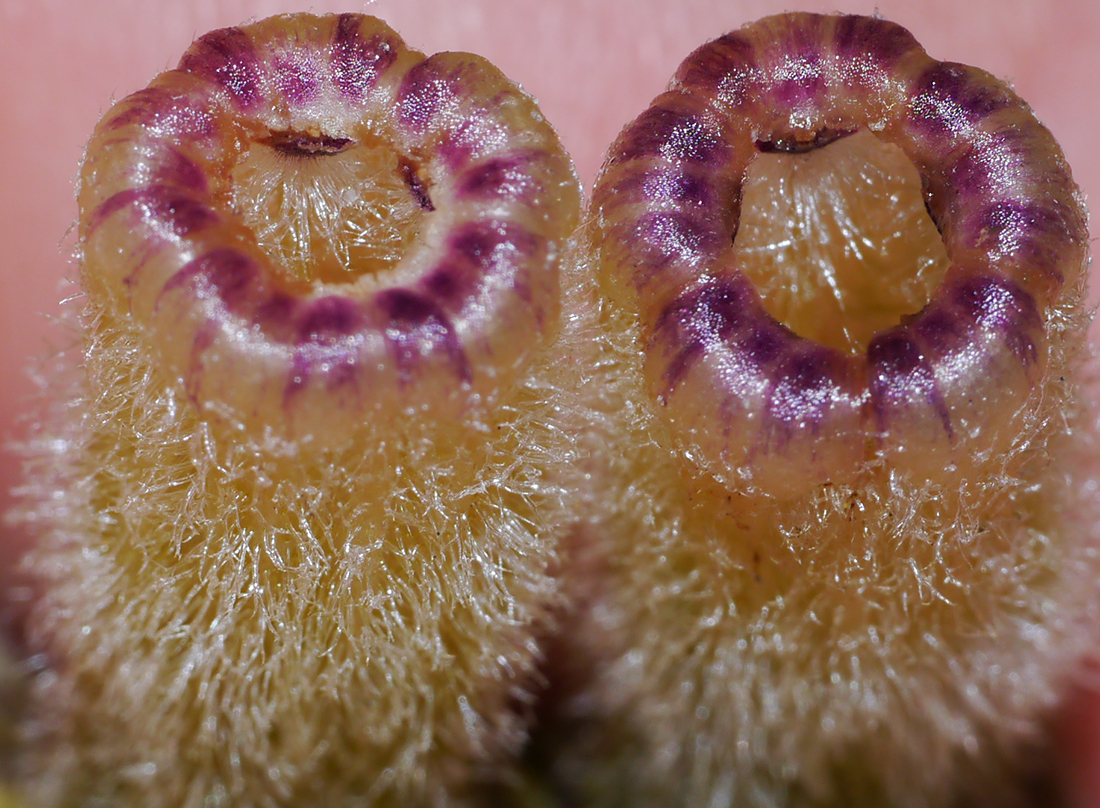
| Figure 13. Ventral (inner) face of anther on left, and dorsal (outer) face on right, of J. aijana. Photo by T. Mione, cultivated plant of Mione et al. 872. Units vertically on right are mm. Two photos, with a slight focus adjustment between, were stacked in photoshop. |
 |
| above, Figure 14. Infructescence (the fruit showing is unripe), pedicels and peduncle of Jaltomata aijana. Units along bottom are mm. Photo by Thomas Mione in Peru, Mione et al. 863. |
Surprisingly, Segundo Leiva G. and T. M. made a single observation (20 May 2015, Mione et al. 863) of fruits of J. aijana with purple pigmentation. This was the first observation of purple pigmentation in the fruits of any species of section Hebecladus (Mione, 1992) and we suspect that the purple fruits we saw on J. aijana were a developmental error and/or atavism.
Observations in Connecticut, Outdoor-grown plants, September 2017.
Bees (unidentified) climb into the tubular corolla of J. aijana and remain in the tube for several seconds before backing out. Flowers harvested and then torn open longitudinally and observed with a dissecting microscope, immediately after this observation, had no nectar. Presumably, the bees that climb into the corolla tube had been consuming the nectar. Mione et al. 872.
ACKNOWLEDGEMENTS
I thank David Spooner for sending his specimens to me, Lindsay T. Kmietek for preparation of figure 1,
Kanchi Natarajan Gandhi for assistance with nomenclature, and Gregory J. Anderson and Gabriel Bernardello for providing an environment conducive to the birth of this project.
Literature Cited